
“Flowers of South-West Europe - a field guide” - de Oleg Polunin e B.E. Smythies
“Revisitas” de regiões esquecidas no tempo - “Plant Hunting Regions” - a partir de uma obra de grande valor para o especialista e amador de botânica como da Natureza em geral.
Por
Horst Engels, Cecilia Sousa, Luísa Diniz, Nicole Engels, José Saraiva, Victor Rito
da
Associação “Trilhos d’Esplendor”
2.13 The Northern Serras of Portugal
2.13 As Serras do Norte de Portugal
2.13.2 Serra da Estrela
|
Folha de Cálculo: Flora da Serra da Estrela
(Lista provisória de plantas vasculares e não-vasculares)
|
Mapas das Serras do Norte de Portugal:
|
2.13.2 Serra da Estrela

Fóssil de folha de Fagus sylvatica (Plioceno) (From Wikipedia)
Subdivisions of the Quaternary System
| |||
Age (Ma)
| |||
0–0.0117
| |||
Pleistocene
|
0.0117–0.126
| ||
0.126–0.781
| |||
0.781–1.806
| |||
1.806–2.588
| |||
older
| |||
In Europe and North America, the Holocene is subdivided into Preboreal, Boreal, Atlantic, Subboreal, and Subatlantic stages of the Blytt-Sernander time scale. There are many regional subdivisions for the Upper or Late Pleistocene; usually these represent locally recognized cold (glacial) and warm (interglacial) periods. The last glacial period ends with the cold Younger Dryas substage.
|
2.13.2d5 - Aspectos glaciários - Filogeografia (Refúgios de Árvores na Península Ibérica)
Falamos no post anterior sobre a filogeografia de Galemys pyrenaica. O estudo de Igea, A. et. al. (2013)[1] que apresentamos no post anterior sobre a filogeografia de Galemys pyrenaicus na Península Ibérica incluiu também modelos sobre a distribuição potential actual e pleistocénica (GLM) desta espécie. Os modelos usam como dados de fundo (background) variáveis climáticas.
No entanto, será interessante de ver qual a vegetação que existe ou existia no tempo da última glaciação nas áreas de potencial distribuição de Galemys pyrenaicus. Esta questão tem interesse porque pode haver uma correlação entre a distribuição desta espécie e outras espécies, também do reino das plantas, que se encontram nos habitats de Galemys pyrenaicus ou que têm preferências climáticas parecidas como esta espécie. São sobretudo as regiões submontanas nos andares meso- e supra-temperadas com florestas caducufólias no norte-oeste da Península Ibérica onde Galemys ocorre e onde se encontra normalmente também floresta caducifólia com Fagus sylvatica e Alnus glutinosa. Mas na Serra da Estrela Fagus sylvatica não existe naturalmente (existe apenas plantado). São as espécies de árvores caducifólias Betula celtiberica e Alnus glutinosa entre outros que são indicativos e autóctones nas zonas de altitude da Serra da Estrela onde Galemys pyrenaicus ocorre.
Aspectos fotográficos de paisagens (Sabugueiro, Rio Alva - Serra da Estrela) e (Covão da Ametade, Rio Zezere - Serra da Estrela) onde ocorre Galemys pyrenaicus

Vale do Rio Alva, Sabugueiro (Serra da Estrela).

Folhagem de outono de castanheiros (Castanea sativa). Sabugueiro, Serra da Estrela

Floresta introduzida ou semi-natural na Serra da Estrela (Sabugueiro)

Rio Alva em Sabugueiro onde foi observado Galemys pyrenaicus em 1971 (Engels, 1972)






A vegetação ribeirinha ainda verde em Julho 2013

O “Covão da Ametade” - com uma represa de água da nascente do Rio Zezere que no entanto secou este ano. Galemys pyrenaicus ocorre segundo (Carvalho, H. 1994) na parte superior do Rio Zezere. Aqui podia ocorrer Galemys pyrenaicus se a represa estivesse com água.
 |
Espécies de bétulas plantadas (Jan Jansen, 2002). Mas ainda existem segundo Jan Jansen algumas manchas com exemplares de Betula celtiberica autóctones na Serra da Estrela.
Também a fauna de macroinvertebrados nos rios onde existe Galemys pyrenaicus pode ser um indicador importante do habitat desta espécie, como por exemplo espécies de plecópteros e triquópteros como de efemerópteros, cuja presença não está apenas ligado à qualidade e características da água, mas também à existência de diversos substratos de materia orgânica e anorgânica.
Será possível demonstrar a existência das mesmas componentes específicas dos habitats actuais também nos refúgios glaciares - ou existem diferenças em relação à composição das espécies?
Como a dinâmica de distribuição geográfica das árvores na Península Ibérica durante o Pleistocénico está já bem estudada, será interessante de comparar à seguir a dinâmica da distribuição geográfica das árvores com os modelos de distribuição geográfica potencial de Galemys pyrenaicus na esperança de encontrar respostas para as perguntas anteriores.

Species-distribution modeling of Galemys pyrenaicus. (A) Potential distribution of G. pyrenaicus as estimated by Maxent for present time. The blacks dots represent occurrence data used for this analysis. Colors indicate probability of presence. (B) Potential distribution of G. pyrenaicus during the Last Glacial Maximum.
Existe uma revisão recente de Arroyo et. al. (2010) [2] sobre a dinâmica de distribuição geográfica de árvores na Península Ibérica durante a glaciação pleistocénica, sobretudo durante a última glaciação Würm, que pode nós dar as pistas necessárias para as questões acima colocadas. Uma lista com os sítios onde foram encontrados restos pleistocénicos de plantas na Península Ibérica encontra-se no review de P. González-Sampériz et al. (2010) [3] sobre a vegetação pleistocénica da Península Ibérica.
.
Podemos dividir as árvores da Península Ibérica em 4 grandes categorias, (a) árvores de folha larga caduca , (b) árvores de folhagem esclerófila perenne, (c) gimnospermas e (d) árvores de folhagem larga perenne.
Para a distribuição geográfica de Galemys pyrenaicus será o grupo (a) árvores de folhagem larga caduca que será de maior interesse porque é o clima temperado e a distribuição geográfica das árvores de folhagem cadúca na Península Ibérica que coincidem com a distribuição geográfica actual da espécie.
Arroyo J. et. al. (2010) escrevem na revisão:
Temperate deciduous forests are mostly distributed along the northernmost fringe of the Iberian Peninsula, in mountain ranges that are under the influence of relatively mild Mediterranean–Atlantic climates, or as gallery forest along water courses. The largest deciduous forests are dominated by white oaks (Quercus spp., subgen. Quercus) and in some regions by beech (Fagus sylvatica L.). Both have been intensively studied within the frame of European-wide research programmes (Petit et al., 2002a,b; Magri et al., 2006). These studies outstand for their integration of population genetic and palaeoecological approaches, and their results are based on an empirical evidence of rarely achieved quality.
Felizmente existe então documentação sobre Fagus sylvatica que é a espécie que mais parece ligada aos biótopes onde se encontra actualmente Galemys pyrenaicus. Eu (Horst Engels) lembro pessoalmente quando acompanhei nos anos setenta Ernst Juckwer, um colega de estudo que na altura estava a fazer a sua tese de doutoramento sobre Galemys pyrenaicus, em duas viagens para o Norte da Espanha (mais especificamente para a Serra de Cameros perto de Logroño) de Ernst Juckwer ter colocado esta hipótese de Galemys pyrenaicus estar ligado troficamente à faja (Fagus sylvatica). Nos locais onde nós apanhamos Galemys pyrenaicus havia sempre Fagus sylvatica em abundância e nos pequenos rios onde a topeira aquática existia formavam-se frequentemente acumulações de detritus e de folhas das fajas. Também lembro-me da presença de larvas de triquópteros e de plecópteros que existiam em abundância por baixo das pedras nestes rios.
Quando se olha para a distribuição geográfica actual de Galemys pyrenaicus e para os seus supostos refúgios pleistocénicos, então uma correlação com a distribuição geográfica pleistocénica de faja (Fagus sylvatica) parece evidente. Pelo menos as áreas de distribuição coincidem em grande parte.
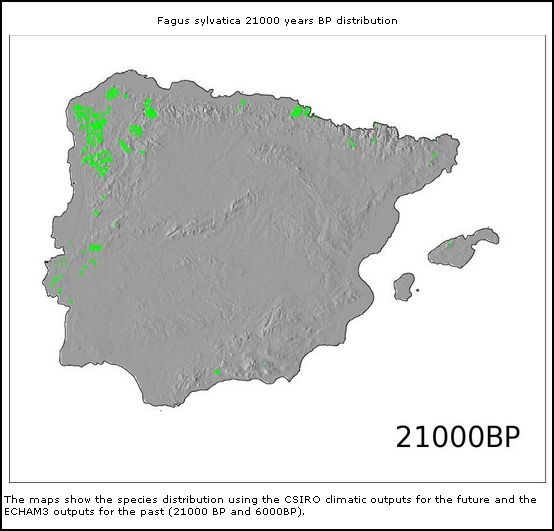 * *
Fagus sylvaticus (21.000BP)
| 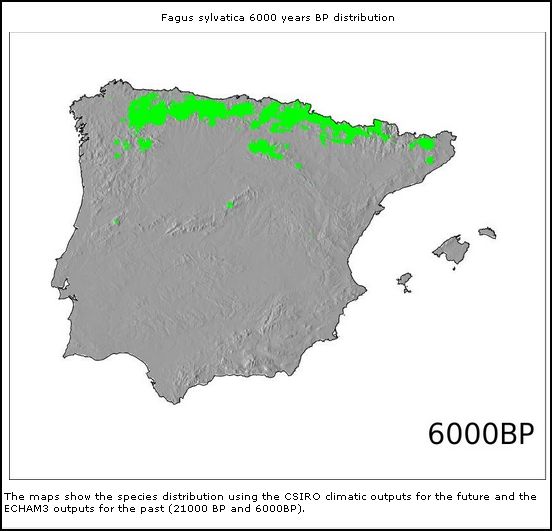 * *
Fagus sylvaticus
(6.000BP)
| 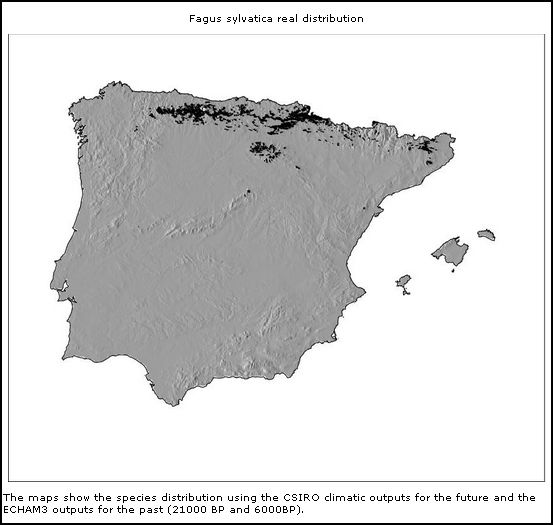 * *
Fagus sylvaticus
(actual)
|
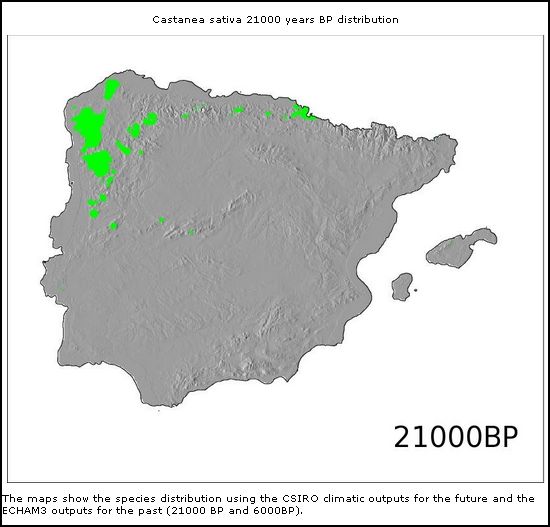 * *
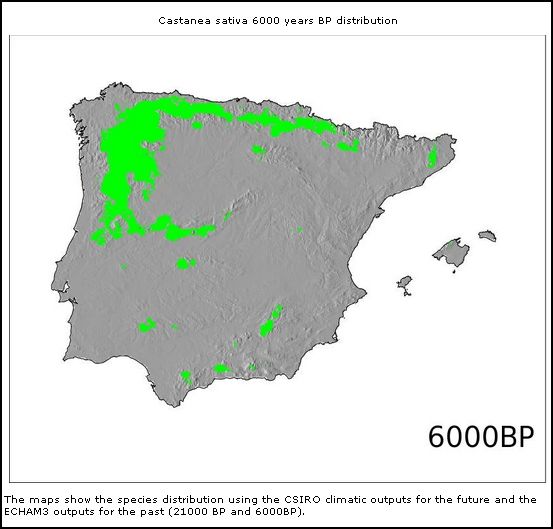
Castanea sativa
(21.000BP)
|  * *
Castanea sativa
(6.000BP)
|  * *
Castanea sativa
(actual)
|
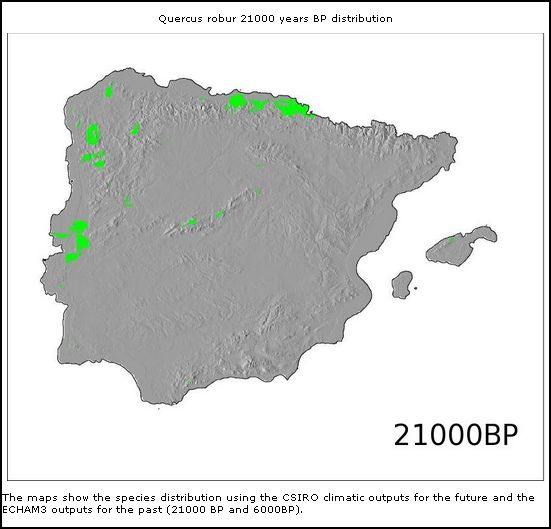 * *
Quercus robur
(21.000BP)
|  * *
Quercus robur
(6.000BP)
| 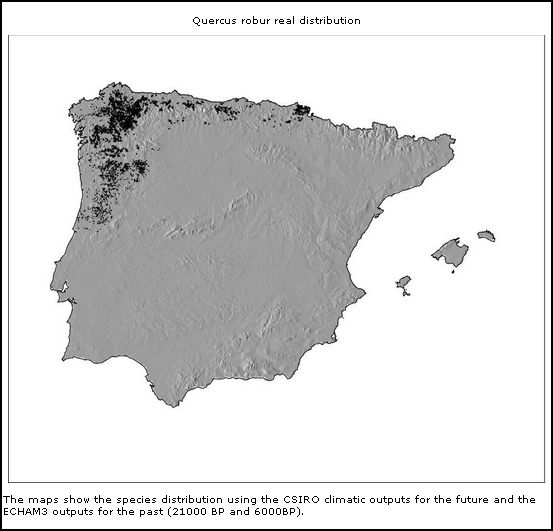 * *
Quercus robur
(actual)
|
Modelos de distribuição de árvores na Península Ibérica
no Pleistoceno Superior e Holocene (From: Marboclim)
Mas também outras espécies de árvores caducifólias como Castanea sativa e Quercus robur demonstram esta correlação aparente de distribuição geográfica entre Galemys e árvores caducifólias.
Se existe na realidade uma dependência trófica indirecta (por exemplo por estados larvares aquáticos de insectos) com as árvores caducifólias acima mencionadas na cadeia alimentar em que Galemys pyrenaicus está inserido ou apenas uma correlação de distribuição geográfica em função do clima tem de ficar ainda em aberto.
Floresta e Fagus sylvatica (From Wikipedia)

Folhas e frutos de Fagus sylvatica (From Wikipedia)
Fóssil de folha de Fagus sylvatica (Plioceno) (From Wikipedia)

Referências da distribuição de Fagus no Pleistoceno.
(From: Jose Maria Postigo Mijarra et. al. 2008)
Mijarra et. al. escrevem (2008):
As regards the beech (Fagus sylvatica L.), although the indigenous origin of this species on the Peninsula is unquestioned, there has on occasions been discussion as to the origin of its populations. Thus, one possibility considered is that following the pre-Holocene extinction of the species on the Iberian Peninsula, this territory might have been re-colonised by populations spreading during the postglacial period from eastern Europe (Balkans) (Huntley and Birks 1983; Demesure et al. 1996), as this was the origin of the population of the taxon on the Peninsula. Hewit (1999) suggests a double origin of the beech populations on the Iberian Peninsula. These were on one hand the tertiary relict populations on the Peninsula and on the other, that from the Balkans in the Holocene re-colonisation. This would confirm the previous theories put forward by other authors (Martinez Atienza and Morla 1992; Costa Tenorio et al. 1997) based on palaeobotanical evidence (Fig. 9). Further studies confirm this idea (Magri et al. 2006).
Encontrou-se Fagus sylvatica na Galiza do Pleistoceno Superior, e nos Pirenéus também do Pleistoceno Inferior. Nas Cantábricas e no Sistema Central Ibérico também há registos do Pleistoceno Médio que coincide bastante bem com os supostos refúgios pleistocénicos de Galemys onde possivelmente se iniciou também a separação subespecífica em duas linhas filogenéticas diferentes. Esta provável existência de Fagus em epócas diferentes do Pleistocénico na Galiza, nas Cantábricas e nos Pirenéus também pode sustentar (pelo menos não contraria) a hipótese que a separação subespecífica de Galemys pode ter se iniciada já antes da última glaciação Würm.
Também, Fagus sylvatica da Península Ibérica tal como Galemys pyrenaicus não faziam parte da recolonização de Europa Central no holocénico a partir dos “refúgios” da Península Ibérica.
Se esperamos no entanto que fortes correlações entre a distribuição de diversas espécies de árvores existissem durante o Pleistoceno na Península Ibérica, ficamos desapontados. A história fitogeográfica das espécies de árvores na Penísula Ibérica parece ser bastante idiosincrática quer dizer cada caso parece ter a sua história individual com refúgios glaciares frequentemente diferentes umas das outras. Está confirmado apenas uma grande tendência de distribuição comum entre a vegetação de árvores caducifólia do clima atlântico temperado do Norte e Norte-Oeste da Península Ibérica e vegetação de árvores xerofíticas perenes do clima mediterrânico do Sul e Sul-Este da Península Ibérica.
Arrojo, J. et. al. (2010) escrevem numa síntese da sua revisão:
...The slow pace of evolution of trees and their abundance in the fossil record render them ideal organisms to throw light on the ‘shallow’ and ‘deep’ phytogeographic history of the Mediterranean Basin (Petit et al., 2005). The case studies reviewed here illustrate that
different species, and populations within species, have responded in a very idiosyncratic manner to past climatic changes. Their population dynamics have been governed by a still poorly understood mixture of manifold causes including the life history, dispersal ability and ecological requirements of species as well as the particular geographical context in which populations perform (Pitelka et al., 1997; Petit et al., 2004). Some common ‘themes’ can however be extracted from the diversity of patterns observed across the Iberian Peninsula.
The most salient feature is the strong and remarkably clear-cut divide between lineages from the Atlantic and the Mediterranean regions of the Peninsula, which is a consistent pattern across species in different genera. This widespread divide is traditionally assigned to the existence of mutually isolated glacial refugia located near each of the two coasts, from which populations would have expanded inland after the LGM until encountering terrains occupied by populations originating in the other refugium. Such a scenario is probably valid for many (or even most) tree species that are currently distributed across the Iberian Peninsula. However, this only explains the maintenance of the east–west divide but not its origin itself. Edaphic preferences of species might be relevant in this regard (Alvarez et al., 2009), as the eastern and the western Iberian Peninsula are characterised by siliceous (acidic) and calcareous (more basic) soils, respectively, that might act as selective filters promoting isolation and differentiation of species and lineages (Blanco et al., 1998). Notwithstanding, the evidence reviewed in this paper allows two more interesting inferences concerning this origin. First, the geographical distribution of genetic lineages and their different genetic variants indicates that the divide probably arose when species entered the Iberian Peninsula from the south and then expanded independently along both coastlines. In contrast, few species entered Iberia apparently from the northeast, and these remained mostly restricted to its Mediterranean sector. The notion of a predominantly southern provenance of currently widespread species is further backed by the fact that several taxa show peculiar and relatively original genetic variants of narrow distribution in populations located in southwestern Spain (e.g. Lumaret et al., 2002; Olalde et al., 2002; see also Fig. 1). Second, the well-developed genetic lineages found within each region indicate that the original formation of the divide probably often antedates the LGM, although not necessarily occurred during the Tertiary.
No entanto, em relação às espécies de carvalho (Quercus spp.) Herran, A. et. al. (1999) demonstram que existem no mínimo 5 regiões de refúgios pleistocénicos para este grupo de espécies na base de haplótipos de ADN encontrados nas espécies de carvalhos na Península Ibérica:
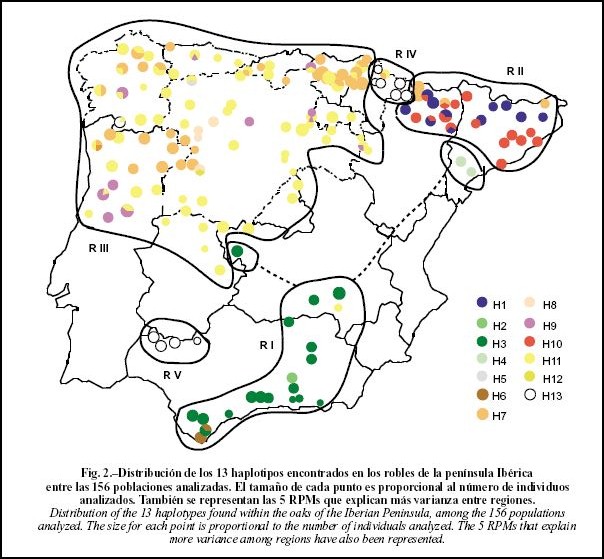
Distibuição de 13 haplotipos de Quercus spp. (Quercus canariensis, Q. faginea, Q.
humilis, Q. pyrenaica, Q. robur y Q. petraea) na Península Ibérica.
E mais tarde, Olalde et. al. (2002) completam ainda este mapa de Herran et. al., (1999) com mais 16 haplotipos novos na construção de uma árvore filogenética que demonstra a existência de 4 linhas filogenéticas diferentes, separando muito bem os carvalhos caducifólios e clima atlântico-temperado do Norte-Oeste dos carvalhos xerofíticos e folhagem perene de clima mediterrânico do Sul e Sul-Este da Península Ibérica como de duas linhas aparte no sul-este provenientes da Itália e dos Balkans.
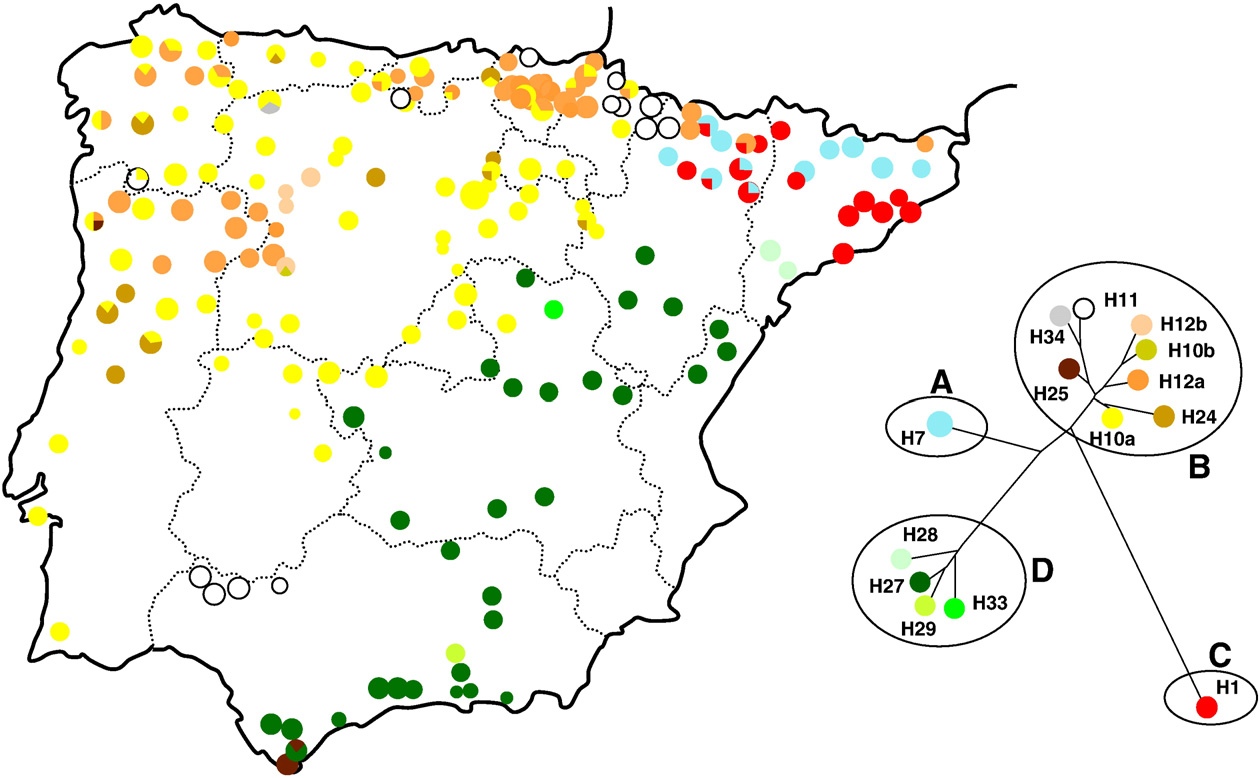
White oaks (Quercus spp.) phylogeography in the Iberian Peninsula (From: Olalde et al., 2002). Note the divide of the major lineages from eastern and western Iberian Peninsula, and the presence of haplotypes of Italian–Balkan origin (H1 and H7, shown in red and light blue colours, respectively) in the northeast. The Gibraltar area harbours haplotypes from both eastern and western lineages, plus one private/endemic haplotype.
Anexo - Maxent (Maximum entropy) modelação da distribuição de espécies
Introduzimos aqui ainda brevemente a ideia da “modelação da distribuição potencial de espécies” usada na modelação potencial actual e pleistocénica de Galemys pyrenaicus por Igea et. al. (2013) e que permite relacionar a distribuição actual das espécies com a sua distribuição potencial tanto actual como em tempos do passado. Esta nova tecnologia da modelação permite complementar bem os trabalhos e filogeografia e em conjunto com a disciplina da filoecologia promete dar alargadas conclusões sobre a biogeografia das espécies.
A técnica de modelação “Maxent - Maximum entropy” usado no trabalho de Igea et. al. (2013) faz uso apenas da presença de espécies actuais e da correlação com outros factores como clima observado em mapas de uma região mais ampla do que as da distribuição actual.
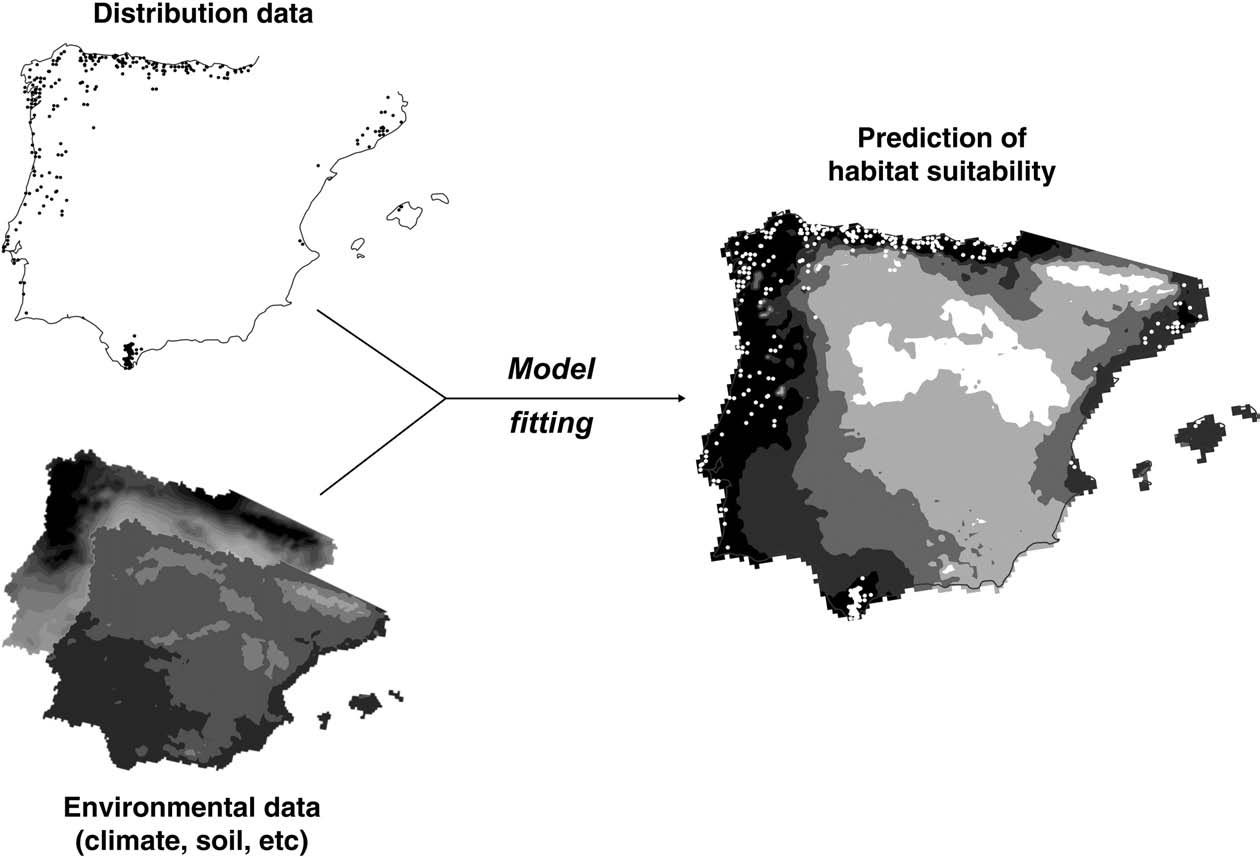
Outline of the workflow to predict species ranges by means of species distribution models. These models relate distribution data with the environmental conditions at sites
where species are present in order to predict suitable areas for the species in past, present or future environmental conditions. Sometimes, data on known absences of the species at
particular sites are also used (see Guisan and Zimmermann, 2000).
4. An introduction to species distribution models (Arrojo, J. et. al. (2010):
In addition to palaeobotanical and phylogeographical data, a new source of information about past distributions of species has recently been added to the biogeographer's toolbox. Species distribution models comprise a broad array of statistical approaches (including generalised additive and linear models, neural networks, regression trees or maximum-entropy algorithms) that relate species distributions to particular environmental variables—predominantly, although not exclusively, related with climate (Guisan and Zimmermann, 2000). The principal aims of such modelling exercises are to gain a better understanding of the factors that shape the geographical distribution of species, and to infer their geographical ranges for different time periods or regions (Pearson and Dawson, 2003; Morin et al., 2007).
There are currently two main approaches to model species distributions (see Guisan and Zimmermann, 2000; Pearson and Dawson, 2003; Guisan and Thuiller, 2005; Kearney and Porter, 2009 for reviews). Mechanistic or process-based models are based on a sound knowledge of the ecology of species and their physiological tolerance to environmental conditions, whereas correlative models seek statistical associations between the current distribution of species and selected environmental variables of potential relevance that can be considered as predictors. Mechanistic models usually require a species-specific collection of long-term field and/or laboratory data, which are often costly and hard to obtain (Kearney and Porter, 2009). In contrast, correlative models are relatively easy to implement due to their phenomenological nature, and their implementation is favoured by the rapidly increasing availability of environmental data with appropriate large-scale geographical coverage and resolution (Graham et al., 2004). Although species distribution models are generally built for particular species or subspecies, they may also be applied to plant functional types (e.g. Hilbert et al., 2007) or other ad hoc groups of organisms. Correlative models have proved to be useful for predicting species distributions across geographical regions and time periods (Araújo et al., 2005; Thuiller et al., 2005; Hijmans and Graham, 2006), even though they are based on some wide-ranging and partly unrealistic biological assumptions (see below). Recent studies have attempted to overcome some of these limitations, e.g. by combining them with spatially explicit population models (Keith et al., 2008) or introducing dispersal processes during range expansion (De Marco et al., 2008). Concomitantly, niche-based species distribution models are increasingly applied in an evolutionary context, to address questions such as niche evolution across phylogenies (Yesson and Culham, 2006; Evans et al., 2009) or the ecological delimitation of lineages (Kozak et al., 2008).
Two kinds of data are required to build a correlative species distribution model (Fig. 3): (1) a list of geo-referenced occurrences of the species (which may be complemented by known absences); and (2) a corresponding dataset of environmental variables (e.g. climate, soil types, and topography). By combining these two data layers one can extract the environmental conditions at localities where the species is present versus those where it is absent (Guisan and Zimmermann, 2000). Once the statistical relationship between species distributions and the environment has been fitted and the predictive ability of the model been verified, it can be used (with due caution) to infer the distribution of species across time and space by projecting its ‘bioclimate envelope’ onto past or future climate scenarios. This extrapolation onto different spatial or temporal realms relies upon a series of assumptions, which have been subject to debate and critiques of the approach (Davis et al., 1998; Hampe, 2004; Dormann, 2007). For instance, the environmental requirements of species are assumed to remain stable over time, and their current geographical distribution is assumed to be environmentally-driven and in equilibrium. The latter may be difficult to justify for some Mediterranean species whose distribution has been largely influenced by human activity, though there are ways to circumvent this limitation (e.g. Latimer et al., 2006). The implications of these assumptions are exacerbated by the fact that the realism and predictive value of model outputs are very difficult to evaluate, because appropriate tests require independent data. Attempts have been made using fossil evidence (Pearman et al., 2008; Rodríguez-Sánchez and Arroyo, 2008), new locations of invasive species in their recently colonised range (e.g., Thuiller et al., 2005), or phylogeographical records (Hugall et al., 2002).
Until recently, most studies using species distribution models have focused on the impacts of modern climate change on future geographic ranges (Guisan and Thuiller, 2005). However, an increasing number of studies is attempting to reconstruct past species distributions (Cheddadi et al., 2006; Martínez-Meyer and Peterson, 2006; Hilbert et al., 2007; Pearman et al., 2008; Rodríguez-Sánchez and Arroyo, 2008; see Nogués-Bravo, 2009 for a review). Most works have focused on the LGM or the mid-Holocene climatic optimum (c. 6 kyr BP), because these periods count with relatively precise and well-established palaeoclimatic reconstructions (Cane et al., 2006). In contrast, the lack of suitable data has until very recently limited analyses of species range dynamics farther back in the past. This is however changing with the advent of palaeoclimate reconstructions for earlier periods, and first modelling exercises (e.g., Rodríguez-Sánchez and Arroyo, 2008) have extended their projections to periods earlier than the Pleistocene (see also Yesson and Culham, 2006 for a combined phylogenetic approach).
Nonetheless, the reliability of palaeodistributions inferred from models fitted to present data is likely to decrease as one moves further back in time, because other confounding processes (e.g. biological interactions or niche evolution) may become increasingly relevant (Pearman et al., 2008). Furthermore, model outputs ought to be understood as estimates of environmental suitability rather than a ‘portrait’ of the actual distributions of species, which may well be shaped by other factors such as interspecific competition or dispersal limitation beyond the fulfilment of climatic or environmental requirements (Davis et al., 1998; Hampe, 2004). In other words, species may have been actually absent from areas inferred to be suitable according to the model. Thus, the hypothetical role of these areas as actual refugia needs to be confirmed by palaeobotanical or phylogeographical data. Additionally, several modelling strategies and palaeoclimatic reconstructions should ideally be used in order to quantify and reduce the uncertainty of the projected palaeodistributions (Nogués-Bravo, 2009). Furthermore, estimates of the uncertainty surrounding the reconstructed palaeodistributions should also be provided, in order to assess the robustness of the inferred distribution scenarios.
How can palaeoecology and distribution modelling inform each other? For instance, palaeoecological data can help test model predictions as well as provide independent evidence to reject or refine proposed palaeodistributions. In turn, species distribution models may be successfully used to identify unknown refugia (e.g. Hilbert et al., 2007; Leroy and Arpe, 2007), particularly of taxa with scant or absent palaeobotanical remains.
|
[1] Javier
Igea, Pere Aymerich, Angel Fernández-González, Jorge González-Esteban,
Asunción Gómez, Rocío Alonso, Joaquim Gosálbez and Jose Castresana:
Phylogeography and postglacial expansion of the endangered semi-aquatic mammal Galemys pyrenaicus. BMC Evolutionary Biology 13:115. 2013.
[2] Francisco Rodríguez-Sánchez, Arndt Hampe, Pedro Jordano, Juan Arroyo: Past tree range dynamics in the Iberian Peninsula inferred through phylogeography and palaeodistribution modelling: A review. Review of Palaeobotany and Palynology 162 (2010) 507–521
[3] P. González-Sampériz et al. Steppes, savannahs, forests and phytodiversity reservoirs during the Pleistocene in the Iberian Peninsula. Review of Palaeobotany and Palynology 162 (2010) 427–457

Sem comentários:
Enviar um comentário