
“Flowers of South-West Europe - a field guide” - de Oleg Polunin e B.E. Smythies
“Revisitas” de regiões esquecidas no tempo - “Plant Hunting Regions” - a partir de uma obra de grande valor para o especialista e amador de botânica como da Natureza em geral.
Por
Horst Engels, Cecilia Sousa, Luísa Diniz, Nicole Engels, José Saraiva, Victor Rito
da
Associação “Trilhos d’Esplendor”
2.13 The Northern Serras of Portugal
2.13 As Serras do Norte de Portugal
2.13.5 Serra do Gerês
|
Bases de Dados:
|
Mapas das Serras do Norte de Portugal e do Gerês:
|
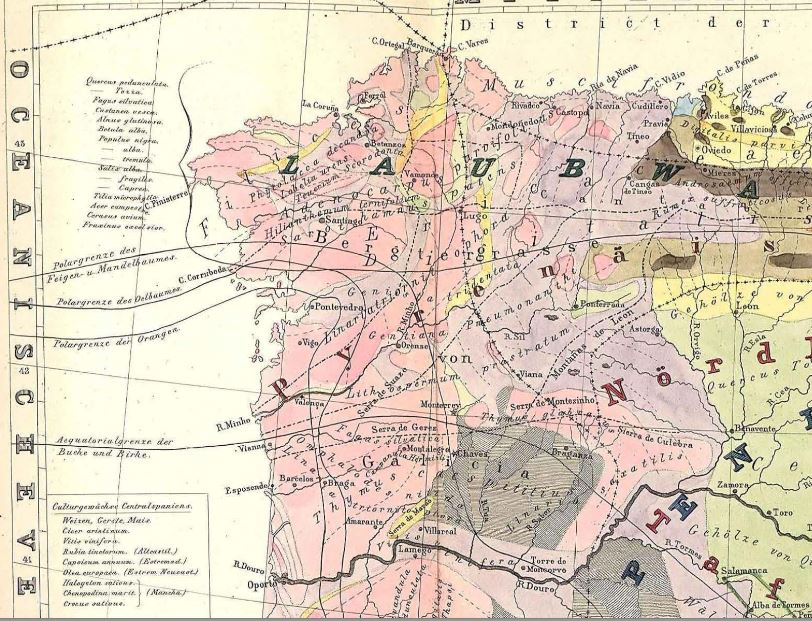
Moritz Willkomm (1852)[1]
Na
Wikipedia encontra-se informação geral sobre aspectos históricos, geográficos,
biológicos, paisagísticos e sociológicos do Parque Nacional da Serra do
Gerês:
A Serra do Gerês (em galego, Serra do Xurés) é a segunda maior elevação de Portugal Continental. O cume atinge 1546 metros de altitude[2] (Pico da Nevosa, na fronteira com a Galiza), segundo folha do Instituto Geográfico do Exército. Faz parte do sistema montanhoso da Peneda-Gerês.
O Parque Nacional da Peneda-Gerês ou conjunto serrano da Peneda-Gerês" , é o único parque nacional de Portugal e situa-se no extremo nordeste do Minho, estendendo-se até Trás-os-Montes, desde as terras da Serra da Peneda até a Serra do Gerês - daí a sua designação -, sendo recortado por dois grandes rios, o Rio Lima e Cávado. Fazendo fronteira com a Galiza, abrangendo os distritos de Braga (concelho de Terras de Bouro), Viana do Castelo (concelho de Melgaço, Arcos de Valdevez e Ponte da Barca) e Vila Real (concelho de Montalegre) numa área total de cerca de 70 290 hectares.[3]

Logótipo do Parque
É
uma das maiores atracções naturais de Portugal, pela rara e
impressionante beleza paisagística e pelo valor ecológico e etnográfico[4] e pela variedade de fauna (corços, garranos, lobos, aves de rapina) e flora (pinheiros, teixos, castanheiros, carvalhos e várias plantas medicinais). Estende-se desde a serra do Gerês, a Sul, passando pela serra da Peneda até a fronteira espanhola.[5]
Inclui trechos da estrada romana que ligava Braga a Astorga, conhecida como Geira.[6] No parque situam-se dois importantes centros de peregrinação, o Santuário de Nossa Senhora da Peneda, réplica do santuário do Bom Jesus de Braga, e o de São Bento da Porta Aberta, local de grande devoção popular.[7]
Localidades
Nas localidades no interior do parque, a vida quotidiana mantém raízes firmes na tradição rural portuguesa. Algumas das de maior valor turístico são:
Destaca-se pelo seu castelo medieval (Castelo de Castro Laboreiro). É famosa pela raça de cães Castro Laboreiro
Destaca-se por possuir o maior aglomerado de espigueiros antigos da Península Ibérica, e, pelo seu castelo do século XIII (Castelo de Lindoso).
Aldeia com grande aglomerado de antigos espigueiros, assentes num afloramento rochoso, com vistas para o vale do Rio Lima.
Esta pitoresca aldeia destaca-se pela arquitectura popular antiga e pelas antigas tradições ainda preservadas.
- O fojo: a alma de (lobo em) Fafião!
Uma
curva aqui, uma curva acolá por entre os penhascos e precipícios
gritantes da magnífica Serra do Gerês, para conseguirmos encontrar a
curiosidade que nos tinham relatado certo dia: o fojo do lobo, uma série
de muros de pedra, construídos na Idade Média, que serviam como
armadilha de lobos.12
- O Vale (encantado) de Albergaria...
[Encantado]
encantar, incantare: in=em; cantare="cantar", aqui com o sentido de
"emitir palavras mágicas". Então encantar será "emitir palavras mágicas e
lançá-las em alguém". Ou, então, "lançar um feitiço em alguém".
Pincães
é um lugar que se destaca pela beleza das cercanias com os seus prados
verdejantes, pelos seus campos de oliveiras e pelo sossego.
Aldeia
encostada à serra, constitui-se como paradigma da sobrevivência, em
tempos remotos, de uma comunidade agro-pastoril. Desse tempo ainda
restam, além dos próprios campos de cultivos ao redor do espaço, o
moinho de cubo e outro de levada, as poças e o gado comunitário. São
frequentes as idas do gado para a serra pastoreado por gente da aldeia.
Paisagem

Panorâmica da Albufeira da Caniçada,
Vila do Gerês, Terras de Bouro
A
natureza e orientação do relevo, as variações de altitude e as
influências atlântica, mediterrânica e continental traduzem-se na
variedade e riqueza do coberto vegetal: matos, carvalhais e pinhais,
bosques de bétula ou vidoeiro, abundante vegetação bordejando as linhas
de água, campos de cultivo e pastagens. As matas do Ramiscal, de
Albergaria, do Cabril, todo o vale superior do rio Homem e a própria
Serra do Gerês são um tipo de paisagem que dificilmente encontra em
Portugal algo de comparável.
Estas
serranias já foram solares do Urso pardo e da Cabra montesa. O Lobo
vagueia num dos seus raros territórios de abrigo. A Águia-real pontifica
no vasto cortejo das aves. Micro-mamíferos vários, caso da
Toupeira-de-água, diversidade de répteis e anfíbios e uma fauna
ictiológica que inclui a Truta e o Salmão enriquecem o quadro zoológico.
É
um momento estranho. Parece que, de repente, ficamos extasiados perante
tanta beleza. No coração deste vale (de Albergaria), diante do silêncio
entrecortado pelo ruído do rio, assumimo-nos encantados perante tamanha
beleza.
História

O passado traduz-se nos castelos de Castro Laboreiro e do Lindoso, monumentos megalíticos e testemunhos da ocupação romana. A geira, o antigo caminho que conduzia os legionários de Bracara Augusta a Astorga, sobrevive num trecho da antiga calçada e nos curiosos marcos miliários.
Curiosos povoados, a arquitectura dos socalcos, paradas de espigueiros,
a frescura dos prados de lima, animam um quadro em que a ruralidade
ainda está presente.
Há também a Ponte da Mizarela, segundo a lenda foi construída pelo demónio, esta antiga ponte românica situa-se na freguesia de Ferral, no concelho de Montalegre.
Nesta ponte travou-se uma importante batalha contra os franceses a
quando das invasões, em que os populares saíram vitoriosos, derrotando
assim o exercito francês.

Passando
à frente e seguindo o trilho, eis que chegámos ao nosso destino. Por
entre as ervas do caminho e as rochas que dificultam a visão, aparece,
com algum ruído tranquilo de água, o Poço Azul...
Actualidade
Actualmente,
o PNPG tem cerca de 240 espécies de fauna vertebrada identificadas no
território e 1100 de flora, além de 500 sítios de interesse histórico e
arqueológico.
Incêndios
De
acordo com dados do Instituto de Conservação da Natureza e das
Florestas, entre 2000 e 2012 registaram-se 1098 incêndios no Parque
Nacional, com uma área ardida de 31901 hectares. Equivale a cerca de 46%
dos 69596 hectares de área total do PNPG mas, como há muitas áreas
sujeitas a mais do que um incêndio naquele período, a área ardida
corresponde a 38% do total .
Ver também
- Serra da Peneda
- Serra Amarela
- Serra do Soajo
- Serra do Gerês
- Cascata do Arado
- Cascata de Leonte
- Cascata da Laja
- Alto do Borrageiro
- Pico da Nevosa
- Portela do Homem
- Mata da Albergaria
Referências
- In: Leonel de Vieira. Portugal Património: Guia - Inventário (em ). Casais de Mem Martins, Rio de Mouro: Círculo de Leitores, 2007. Capítulo G02. p. 10. vol. II. ISBN 978-972-42-3908-8.
- Parque Nacional Peneda-Gerês www.cm-melgaco.pt. Visitado em 13 de Janeiro de 2010.
- Parque Gerês/Xurés é reserva da biosfera. Visitado em 27 de Maio de 2009.
- [portal.icnb.pt - CARACTERIZACAO PATRIMONIO (POAP) - Revisão do plano de ordenamento do parque nacional da Peneda-Gerês]
- À Descoberta do Parque Nacional Peneda-Gerês www.destinoseviagens.com. Visitado em 13 de Janeiro de 2010.
- A Geira na Serra do Gerês geira.cm-terrasdebouro.pt. Visitado em 13 de Janeiro de 2010.
- patrimonio cultural pnpg - ICNF portal.icnf.pt. Visitado em 13 de Janeiro de 2014.
- Castro Laboreiro www.cm-melgaco.pt. Visitado em 13 de Janeiro de 2010.
- Castelo alfarrabio.di.uminho.pt. Visitado em 13 de Janeiro de 2010.
- Serra do Geres - PNPG - Turismo - Localidades www.serradogeres.com. Visitado em 13 de Janeiro de 2010.
- Parque Nacional da Peneda-Gerês, Descubra Portugal www.descubraportugal.com.pt. Visitado em 13 de Janeiro de 2010.
- O Fojo do Lobo - Fafião www.serra-do-geres.com. Visitado em 22 de Junho de 2011.
- O Vale [encantado de Albergaria...] www.serra-do-geres.com. Visitado em 22 de Junho de 2011.
- Pincães www.serra-do-geres.com. Visitado em 14 de setembro de 2014.
- Xertelo www.serra-do-geres.com. Visitado em 14 de setembro de 2014.
- flora pnpg - ICNF portal.icnf.pt. Visitado em 13 de Janeiro de 2010.
- Parque Nacional de Peneda-Gerês www2.ufp.pt. Visitado em 13 de Janeiro de 2010.
- Paisagem do Vale de Albergaria-Gerês www.serra-do-geres.com. Visitado em 22 de Junho de 2011.
- actividades humanas pnpg - ICNF portal.icnf.pt. Visitado em 13 de Janeiro de 2014.
- Poço Azul - a Epifania... www.serra-do-geres.com. Visitado em 13 de Janeiro de 2010.
- Centenas de fotografias ilustram em livro os 40 anos do Parque Nacional Peneda-Gerês.
- 38% da Peneda-Gerês ardeu em 12 anos.
Uma
excelente caracterização da biodiversidade, geodiversidade, do
património histórico-arqueológico, da paisagem e das estruturas físicas e
socio-económicas do Parque Nacional da Serra do Gerês, incluindo cartas
e mapas, encontra-se nos Relatórios sobre o Plano de Ordenamento (POPNPG)[8] do ICNF:
- Lista dos Relatórios da Fase de Caracterização (POPNPG)
- Lista das Cartas da Fase de Caracterização (POPNPG)
Bioclima, Biogeografia, Vegetação actual e potential
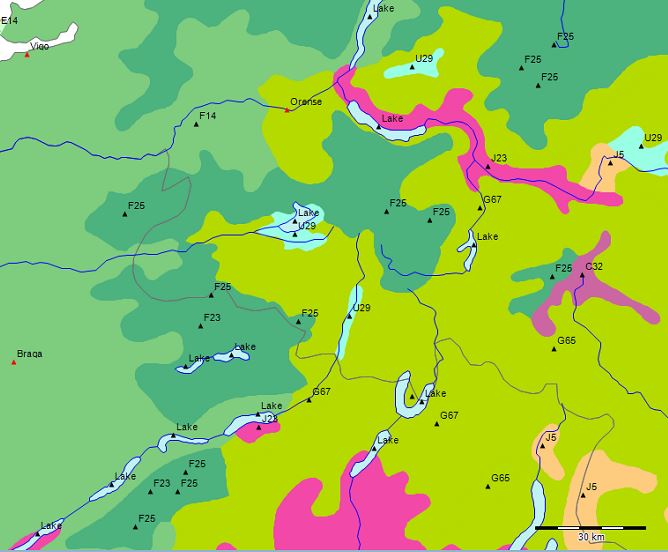
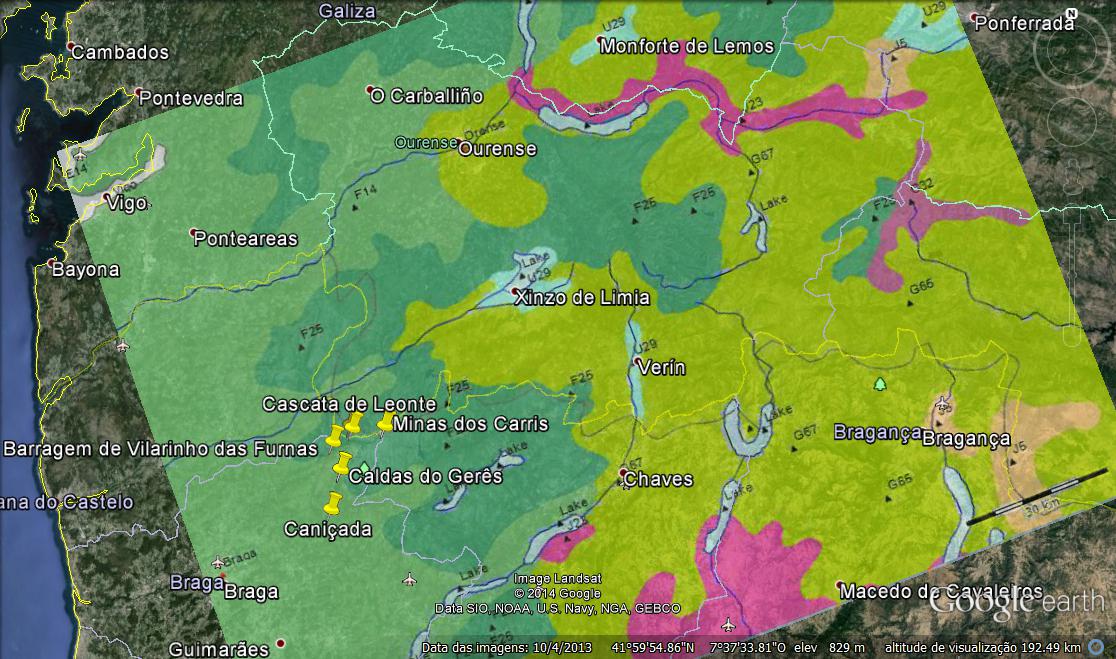
Extrato do ‘Mapa da Vegetação Natural Potencial da Europa’ (U. Bohn et. al. 2000) - Sobreposição da imagem no Google-Earth
A floresta do Parque Nacional da Serra do Gerês é uma floresta composta por carvalhos de folhas largas como Quercus robur, Quercus pyrenaica e de gimnospermas como Pinus sylvestris e de Taxus baccata, mas encontram-se também betuláceas e espécies como Castanea sativa nos tipos de florestas características para a região.
No Mapa da Vegetação Natural e Potencial da Europa (U. Bohn et. al. 2000) está indicada a presença dos tipos F23 (Galician-north Lusitanian oak forests (Quercus robur, Q. pyrenaica) with Betula pubescens subsp. celtiberica, Cytisus striatus, Dryopteris aemula, Anemone trifolia subsp. albida, Omphalodes nitida) e F25 (Orocantabrian-northwest Iberian altimontane birch-sessile oak forests (Quercus petraea, Betula pubescens subsp. celtiberica) and birch forests (Betula pubescens subsp. celtiberica) with Saxifraga spathularis, Crepis lampsanoides, Doronicum carpetanum, Poa chaixii, Luzula sylvatica subsp. henriquesii) da formação das florestas caducas de folhas largas e mixtas com coníferos.
F - Mesophytic deciduous broad-leaved and mixed coniferous-broad-leaved forests
1 - Species-poor acidophilous oak and mixed oak forests (Quercus robur, Q. petraea, Q. pyrenaica, Pinus sylvestris, Betula pendula, B. pubescens, B. pubescens subsp. celtiberica, Castanea sativa)
1.3 - Montane and altimontane types

F23 - Galician-north Lusitanian oak forests (Quercus robur, Q. pyrenaica) with Betula pubescens subsp. celtiberica, Cytisus striatus, Dryopteris aemula, Anemone trifolia subsp. albida, Omphalodes nitida
F24 - Orocantabrian-Euscaldian oak forests (Quercus pyrenaica, partly Q. robur) with Pyrus cordata, Linaria triornithophora, Luzula sylvatica subsp. henriquesii, Arenaria montana, Anemone trifolia subsp. albida, Crepis lampsanoides
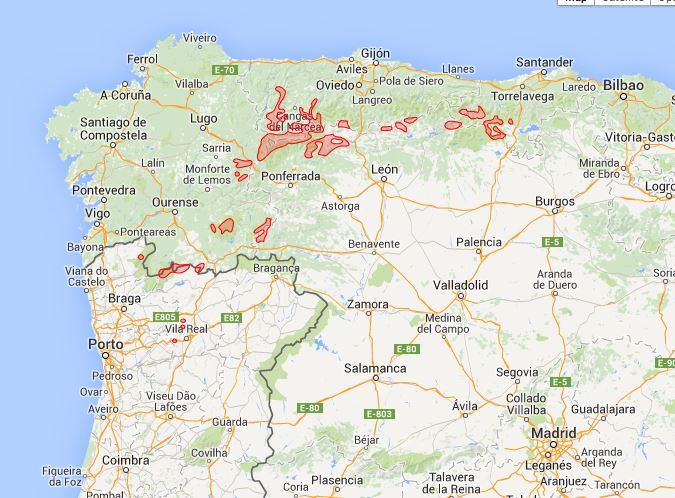
F25 - Orocantabrian-northwest Iberian altimontane birch-sessile oak forests (Quercus petraea, Betula pubescens subsp. celtiberica) and birch forests (Betula pubescens subsp. celtiberica) with Saxifraga spathularis, Crepis lampsanoides, Doronicum carpetanum, Poa chaixii, Luzula sylvatica subsp. henriquesii

F26 - East Pyrenean sessile oak forests (Quercus petraea) with Erica arborea, Daphne laureola, Luzula forsteri, L. sylvatica, Hypericum androsaemum, Galium rotundifolium, Conopodium majus
|
F.1 Species-poor oak and mixed oak forests (Quercus robur, Q. petraea, Q. pyrenaica, Pinus sylvestris, Betula pendula, B. pubescens, B. pubescens subsp. celtiberica, Castanea sativa)
Characterisation and typological delimitation; geographical distribution (J. Cross, J. Pallas)
Acidophilous
oak and mixed oak forests are characterised by the dominance of oaks,
usually Quercus robur and Q. petraea, in the tree layer and of
acidophytes of a Middle European[11] or
temperate Eurasian distribution in the lower layers. The shrub and herb
layers are usually well developed, but are relatively species-poor
compared to deciduous forests on more base-rich sites or in climatically
more favourable areas.
These oak forests extend within the temperate zone of Europe from the Atlantic coast to western Russia (Map 12). The southern boundary locally borders on to montane regions of the submeridional zone. The entire distribution area resembles a fragmented, narrow-angled triangle whose base extends from northern Portugal (approximately 41° N) along the Atlantic coast via Ireland to Scotland (approximately 58° N). The tip of the triangle lies north-east of Kiev (approximately 53° N, 34° E), with isolated outposts as far east as the Volga at Kasan. The northern boundary runs through Scotland, southern Scandinavia, Lithuania and Belarus, and the southern boundary through northern Portugal, northern Spain, the south of France, northern Italy, Croatia, Bosnia-Herzegovina, Romania, Ukraine and southern Russia. Southern outliers of these forests extend into the submeridional zone.
The
soils are characteristically permeable or intermittently moist,
oligotrophic, acidic, often sandy and sometimes shallow or rocky.
As a result of competition from beech, acidophilous oak forests within the natural distribution area of beech, particularly in the sub-Atlantic and Central European floristic provinces, are restricted to intermittently moist soils or very shallow soils on slopes that are too dry for the survival of beech seedlings. The naturalness of acidophilous oak forests is partly a matter of speculation, since in many places coppicing and coppice with standards have favoured oak in the past.
The majority of near-natural stands are scattered and relatively small. Extensive areas occur principally outside the range of beech, i.e. in western Cantabria and Galicia, south-west France, Ireland and Great Britain, Poland, Belarus and northern Ukraine.
The
range of the acidophilous mixed oak forests is not as extensive as that
of the summer-green broadleaved forests in general, because there are
additional factors restricting its distribution:
- In the montane belt of Central Europe and the submeridional zone, beech forests dominate
- In the north and north-east of its range, competition from coniferous forests (Formation D) is the limiting factor. On acidic oligotrophic sites in the east, the deciduous forest formation is less well developed than coniferous stands in comparison with more base-rich sites (cf. MONK 1966, quoted from JÄGER 1969: 401). As a result, pure broadleaved forests are only encountered here on base-rich sites.
- In the east, edaphic and climatic factors (relative lack of precipitation, warmer summers, more nutrient-rich soils) prevent further development on acidic soils. For this reason the eastern border of the acidophilous deciduous forests runs further to the west than the limit of the broadleaved forest formation in general.
- In the temperate zone, acidophilous mixed oak forests favour the lowland to colline altitudinal belts. In the submeridional zone on the other hand, at the southern limit of their range, they are usually associated with the submontane-montane altitudinal belt. Their characteristic species combination gradually changes towards the south and thermophilous sub-Mediterranean oak forests eventually become dominant (Formation G).
Stand structure and physiognomy (J. Cross)
The
natural structure of these forests is often unclear as the activities
of man and his grazing animals over many centuries still have an
influence today. Near-natural stands consist of five to six layers, i.e.
a first and second tree layer, a shrub layer, a dwarf shrub layer, a
herb layer and a moss layer. In some stands, however, only three layers
may be present.
The
tree layer typically has a 60-90 % canopy cover and reaches a height of
15-25 (35) m. On very shallow, dry soils or in very exposed locations
the stand height may be less than 5 m, while on deep soils it can exceed
30 m. The tree layer is dominated by acid-tolerant oaks (Quercus robur, Q. petraea). Pioneer trees such as birches (Betula pendula, B. pubescens)
play an important role in the regeneration phase on different sites,
particularly in the west, and they persist in the tree layer in mature
stands particularly on intermittently moist sites. In the east, pine and
locally also spruce, can occur admixed with the oak to form the tree
layer. The cover and vitality of the shrub layer depends on the density
of the tree layer as well as the nutrient balance, water supply and
humidity of the site. Members of the Ericaceae or brooms (the latter
particularly in more southern areas) often form a dwarf shrub layer and
in the northern temperate areas of Europe the frequent dominance of
dwarf shrubs lend the forests a boreal character. The herb layer is
usually composed of hemicryptophytes, primarily grasses, woodrushes and
sedges. Geophytes are generally rare, but ferns characteristically play
an important role, particularly in the Atlantic and sub-Atlantic
regions. Pteridium aquilinum can form an additional seasonal layer in
some areas, dense stands of this species displacing almost every other
herb species. The percentage cover and number of species in the moss
layer also vary considerably. In the hyperoceanic west, where epiphytic
communities are a striking feature of the forests, cover can be
particularly high. Cryptogam-rich communities also occur on the
north-west European mainland on the edge of poor, dry forest stands,
where the shrub and herb layer is only poorly developed because of
competition from the tree layer.
Floristic composition (J. Pallas)
European
acidophilous mixed oak forests are characterised by a tree layer
dominated by oaks and an undergrowth in which acidophilous species
prevail. Species of the temperate Middle European floristic region
constitute the greater part of the species composition. In the north and
east of the area, the proportion of Eurasian temperate and Eurasian
boreal taxa increases. In the southern temperate regions of Europe,
numerous sub-Mediterranean species occur in the community, while in the
south-west of the area even some Mediterranean species are present (see Table 12).
Apart from the two Middle European oak species (Quercus robur and Q. petraea) widespread tree species include the frequently intermixed beech (Fagus sylvatica), as well as the Eurasian species Betula pendula and Populus tremula. Within the shrub layer the Eurasian species Sorbus aucuparia, Frangula alnus (almost absent within these forests in the British Isles) and Juniperus communis, as well as elements of the Middle European flora, such as Corylus avellana and to a lesser extent Crataegus monogyna, number among the most frequent species. Frequent and widespread species of the herb layer (see Table 12) include the acidophytes Melampyrum pratense, Pteridium aquilinum, Solidago virgaurea, Agrostis capillaris, Anthoxanthum odoratum, Calluna vulgaris, Deschampsia flexuosa, Molinia caerulea, Holcus mollis, Veronica officinalis, Carex pilulifera, Viola riviniana, Potentilla erecta, Lathyrus linifolius and Danthonia decumbens. The mosses, which are mostly acidophytes, have a circumpolar distribution. Polytrichum formosum, Dicranum scoparium, Hypnum cupressiforme, Leucobryum glaucum, Pleurozium schreberi and Scleropodium purum are
the most frequent species (gaps in the Iberian mapping units may be due
to incomplete records of cryptogams in the vegetation surveys).
Representatives of the submeridional zone (in Europe these are species of the sub-Mediterranean sub-region, and to a lesser extent also of the Pontic-south Siberian floristic region), are primarily to be found within southern temperate, and especially sub-Mediterranean, mapping units. Castanea sativa, Sorbus torminalis, Pyrus pyraster, Hieracium sabaudum and Festuca heterophylla are characteristic. In the northern temperate areas of Europe, sub-Mediterranean species are almost completely absent from these forests in the west, while in the east they become more numerous, presumably due to the more marked summer warmth.
The most striking floristic divide runs between the sub-Atlantic and the Central European province. In the Atlantic province, A t l a n t i c species such as Erica cinerea, Hyacinthoides nonscripta and Ceratocapnos claviculata, as well as Atlantic-sub-Atlantic species, are well represented.
In the sub-Atlantic province many A t l a n t i c - s u b - A t l a n t i c species are still encountered, e.g. Teucrium scorodonia, Hypericum pulchrum, Blechnum spicant, Luzula sylvatica, Galium saxatile, Lonicera periclymenum and Hedera helix. Lianas seem to develop their greatest vitality (within the acidophilous oak forests) in the Atlantic areas (Lonicera periclymenum, Hedera helix). In the south Atlantic oak forests, the Mediterranean-Atlantic species Tamus communis and Rubia peregrina also occur.
The great majority of A t l a n t i c species (DT 8.1., 8.1.S in Table 12) belong to the t h e r m a l - o c e a n i c group of species from the western side of the continents in the northern hemisphere (cf. JÄGER 1968). It is therefore no surprise that the role of these species increases continuously within the mapping units in the direction of their climatic optimum in south-west Eurasia.
Ulex europaeus, character species of distribution type 8.1. (lusit-atl), is a typical thermal-Atlantic species. It displays a tendency for a southern Atlantic distribution, which also characterises other species with similar requirements. Frequently, species of this group are absent from the north-west European mainland. Some A t l a n t i c - s u b - A t l a n t i c species within the acidophilous oak forests (DT 8.2., 8.5., 8(1).6.) appear in the northern temperate-euoceanic regions of Europe only in the British Isles and are absent from the European mainland.
Some A t l a n t i c species occur only in the southern Atlantic subprovince, sometimes with outposts in the Meridio-Atlantic province group, e.g. Pyrus cordata, Pseudarrhenatherum longifolium, Daboecia cantabrica, Potentilla montana etc.
These species are grouped together in the newly established Pseudarrhenatherum longifolium distribution type 8.1a. in Table 12 (north lusit-south atl, cf. MEUSEL et al. in 1965, map 44d). The Iberian acidophilous oak forests within the 8.1a. area display an increasing south Atlantic character, in addition to further southern Atlantic species, endemics, such as Omphalodes nitida, Crepis lampsanoides, Aquilegia vulgaris subsp. dichroa, occur here. These taxa are also assigned to the DT 8.1a. The greatest number of Atlantic species occur in the north-west Iberian unit F14.
Another group of species also inhabits parts of the south Atlantic subprovince, but in Iberia and to the south it occurs not only in Atlantic areas, but also in the less oceanic continental interior, where it is characterised by a preference for the montane altitudinal belt. Unlike the DT 8.1a. the greater part of the range of these species is in the s u b m e r i d i o n a l zone. Quercus pyrenaica and Arenaria montana can be regarded as character-species of this distribution type (JALAS & SUOMINEN (Ed.) (1976, 1983) map 305 and 688). Their area corresponds to the western segment of the western sub-Mediterranean DT 5.6. Their distribution type is therefore newly established as the Quercus pyrenaica type 5.6a.: (west med)-west submed//mo-(south atl). With exclusively mountain plants, 6.1a. is also possible. Again, some species, particularly Iberian endemics, only
inhabit parts of this 5.6a.-range.
Additional s u b - M e d i t e r r a n e a n - o c e a n i c species such as Luzula forsteri, Euphorbia amygdaloides, E. dulcis and Polystichum setiferum are likewise concentrated in the southern Atlantic units, but can extend into the central (F17, F21) and the eastern sub-Mediterranean province groups. A frequent distribution type is 5.7.
M e d i t e r r a n e a n species (DTG 1.), such as Quercus suber, Pinus pinaster, Erica arborea, Arbutus unedo, Tamus communis, Rubia peregrina, Ruscus aculeatus, Asphodelus albus, Hypericum androsaemum and Asplenium onopteris within the mapping units considered here only occur in the south-west within the south Atlantic areas, in particular in the lowlandcolline units. F14 contains the greatest relative proportion of these species, followed by F7 and F15. The Mediterranean-Atlantic distribution type 1.10. is frequent, i.e. most species are markedly thermal-oceanic.
In contrast, in the north of the Atlantic province, there are only a few vascular plant species in the table that thrive as well, or even better, than they do in the south and in the Meridio-Atlantic province group, within the acidophilous oak forests. These include Galium saxatile, Blechnum spicant, Luzula sylvatica, Oreopteris limbosperma, Dryopteris aemula, and possibly also Ilex aquifolium and Erica tetralix. Many of these species are montane plants in the eastern part of the sub-Atlantic province.
Most representatives of the Mnium hornum moss
group can be considered as northern temperate-Atlantic-sub-Atlantic. On
particularly poor sites in north-western regions, bryophytes such as Lophocolea heterophylla, Plagiothecium laetum and Aulacomnium androgynum, which grow on raw humus, are characteristic.
The acidophilous oak woods of western Ireland, and to a lesser extent western Britain, are particularly noteworthy. There, h y g r i c h y p e r o c e a n i c, desiccation-sensitive species of cryptogams occur in an abundance not encountered elsewhere in acidophilous oak forests. In the Blechno-Quercetum petraeae KELLY (1981), the average number of species is 57.6.
Spermatophytes constitute about 34 % while cryptogams constitute 66 %. Unit F1 represents an extremely hygric hyperoceanic unit within these oak forests, and is analogous to the extremely thermal-hyperoceanic unit F14 in Iberia. Ferns, such as Hymenophyllum tunbrigense, H. wilsonii and Dryopteris aemula, wider distributed oceanic bryophytes such as Lepidozia reptans and many extremely Atlantic bryophytes such as Dicranum scottianum are characteristic, but also the liverworts grouped around Adelanthus decipiens are characteristic. These species also occur in the laurel forests of the Macaronesian floristic sub-region (azor-canar-mad).
All oceanic species groups characteristic of the Atlantic and sub-Atlantic units are absent from the two eastern provinces of the Middle European floristic region. The mapping units there are characterised more by an increase in b o r e a l and E u r a s i a n t e m p e r a t e taxa, which begin to appear in the north-western temperate areas. Representatives of this species group include Pinus sylvestris, Picea abies, Vaccinium myrtillus, Luzula pilosa, Maianthemum bifolium, Vaccinium vitis-idaea, Trientalis europaea, Rubus saxatilis, Pyrola rotundifolia, Calamagrostis arundinacea and Orthilia secunda. Many of these are coniferous forest elements with c o n t i n e n t a l distribution types (DT 10.3., 10.8., 11.3.). The east European character also becomes more pronounced with the appearance of a range of species of the summer-green broadleaved forests, which are absent from the Atlantic province. The species of the s u b c o n t i n e n t a l Asarum type 8.11., with centres of distribution in the east of the Middle European region, are missing in the Ulex range 8.1.; further, species within the subcontinental 8.13. and 8.14. distribution types should also be grouped here (see Table 12 p. 5). These include Acer platanoides, Euonymus verrucosa, Campanula persicifolia, Carex montana, Peucedanum oreoselinum, Potentilla alba and numerous other species. These species groups play an important role also in the Illyrian F21, located in the eastern part of the central sub-Mediterranean province group, while the Insubrian F17, also in the central sub-Mediterranean, is characterised by the coexistence of oceanic and subcontinental species. Amongst the cryptogams, the species group centred around Dicranum polysetum, differentiates the east European units.
In addition to the generally distributed sub-Mediterranean species already mentioned, the Central European and Sarmatian mapping units, especially the southern Central European F20 and the Illyrian F21, are distinguished by the presence of characteristic eastern sub-Mediterranean species with ranges extending into eastern Middle Europe, e.g. Genista tinctoria, Cytisus nigricans, Genista germanica and others. Nevertheless, a number of representatives of the continental DT 5.9. such as Vincetoxicum hirundinaria or Tanacetum corymbosum can encroach far to the west, particularly into the xerothermic forms of F17 or into the xerothermic sub-Atlantic F18.
The special phytogeographic role of the s u b m e r i d i o n a l zone is underlined by the distribution of oak species in the acidophilous oak forests. Quercus robur and Q. petraea are the stand-forming species in the northern temperate areas of the distribution range. In the southern temperate to sub-Mediterranean areas other oak species occur, such as Q. pyrenaica, Q. canariensis and Q. suber on the Iberian Peninsula or Q. dalechampii, Q. polycarpa, Q. cerris and Q. frainetto in south-east Europe.
…
Mapping units on intermittently moist sites (F3, F9, F22) are characterised by the more frequent occurrence of moisture tolerant species, such as Molinia caerulea, Deschampsia caespitosa, Agrostis stolonifera, Carex nigra and Lysimachia vulgaris. For western units, Myrica gale, Erica tetralix, Narthecium ossifragum and Sphagnum species are characteristic and for Central European-Sarmatian units, Molinia arundinacea and Carex brizoides are typical.
Finally, oreophytes differentiate the montane mapping units. Here, western sub-Mediterranean mountain plants play a role, particularly in F25, while the central sub-Mediterranean F17 is characterised by alpine elements (particularly Luzula nivea and Phyteuma betonicifolium).
Position in the phytosociological system (syntaxa) (J. Pallas)
Depending on point of view the order of the acidophilous oak forests, Quercetalia roboris Tüxen 1931, is placed in the class Querco-Fagetea or, as originally, in an independent class, the Quercetea robori-petraeae Braun-Blanquet et Tüxen 1943.
The reasons underlying these different views relate to the geographical origin of the authors, since, as can be seen in Table 12, the order Quercetalia contains a higher proportion of more demanding deciduous forest species in the southern temperate and submeridional/montane areas of Europe than it does in the north-west European temperate lowlands.
The Quercetalia roboris Tüxen 1931 includes phytogeographically heterogeneous associations, which are characterised by the dominance of both acid indicators and acid-tolerant oaks. The combination of both features is the actual characteristic of the order.
Compared to the Atlantic units that contain numerous species characteristic of extremely oceanic regions (Ireland, north-west Iberia), the sub-Atlantic units behave as marginal syntaxa without any of their own character species. On the other hand the boundary between the sub-Atlantic and Central European province is quite clear, being marked by the loss of Atlantic-sub-Atlantic species. Nevertheless, compared to the Sarmatian units with their prominent share of Eurasian or circum boreal coniferous forest, temperate Eurasian and east European species, as well as their better trophic conditions, the Central European units behave only as marginal syntaxa.
In
principle, only the oak forests of the two main centres of diversity
(atl + submed) are positively floristically characterised by the
presence of acidophytes of a European distribution. From a pan-European
perspective, synsystematic problems arise as a result of the
climatically-determined disappearance of Atlantic and
Atlantic-sub-Atlantic species towards the east. The greater species
diversity in the submeridional zone compared to the temperate zone also
has to be considered (cf. MEUSEL & JÄGER 1989).
In Table 12 an attempt has been made to compare as many units of the geographically widespread order Quercetalia roboris as possible. The following review represents a revision and extension of the proposals previously published by the author (PALLAS 1996, 2000). The division is now based exclusively on chorological criteria. Habitat differences (intermittently moist units) are classified into sub-units according to geographic criteria.
The special position of the extremely oceanic oak forests should be clearly indicated by establishing separate alliances. These units represent centres of diversity in their own right. The Hymenophyllo-Quercion encompasses the Irish hygric hyperoceanic oak forests and the Quercion robori-pyrenaicae constitutes the thermal hyperoceanic oak forests in north-west Iberia. The remaining southern temperate acidophilous oak forests of the Atlantic and sub-Atlantic province belong to the Quercion roboris, and the remaining northern temperate oak forests of the Atlantic and sub-Atlantic province can be grouped in the Molinio-Quercion. In the same way the oak forests of the Central European and Sarmatian province can be grouped into a northern temperate alliance, the Vaccinio-Quercion petraeae and a southern temperate alliance, the Agrostio-Quercion. The central and eastern sub-Mediterranean acidophilous oak forests belong to the Castaneo-Quercion.
...
2. Quercion robori-pyrenaicae (Braun-Blanquet et al. in P. Silva et al. 1950 corr. Br.-Bl. et al. 1956) Rivas-Martínez 1975:
(submeridional-)southern temperate, southern Atlantic, northern
lusit-galic-cant, thermal hyperoceanic (contains Iberian endemic taxa
with the distribution types 8.1a. and 5.6a., see Table 12)
Original form of the name: Quercion roboris broteroanae Braun-Blanquet, Pinto da Silva, Rozeira & Fontes in Pinto da Silva, Rozeira & Fontes 1950, Agronomia Lusitana 12(3): 435. Quercion occidentale Br.-Bl. et al. 1956 = Quercion roboris broteroanae (Braun-Blanquet et al. 1956, Agronomia Lusitana 18(3): 173).
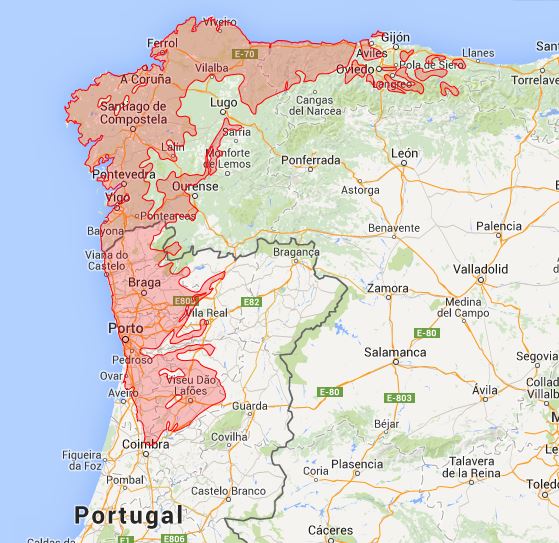
F14 Rusco aculeati-Quercetum roboris Braun-Blanquet, P. Silva et Rozeira 1956; Blechno spicant-Quercetum roboris Tüxen et Oberdorfer 1958.
|
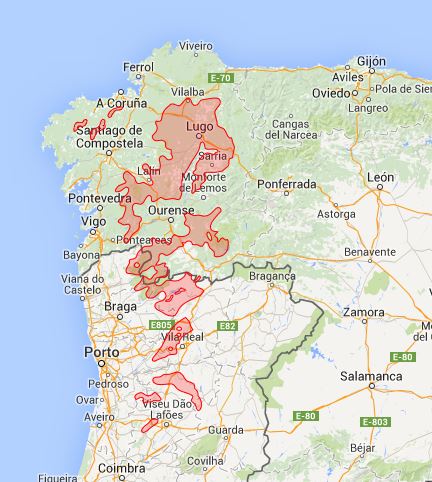
F23 Myrtillo-Quercetum roboris Braun-Blanquet, P. Silva, Rozeira et Fontes in P. Silva, Rozeira et Fontes 1950 (holotype!).
|
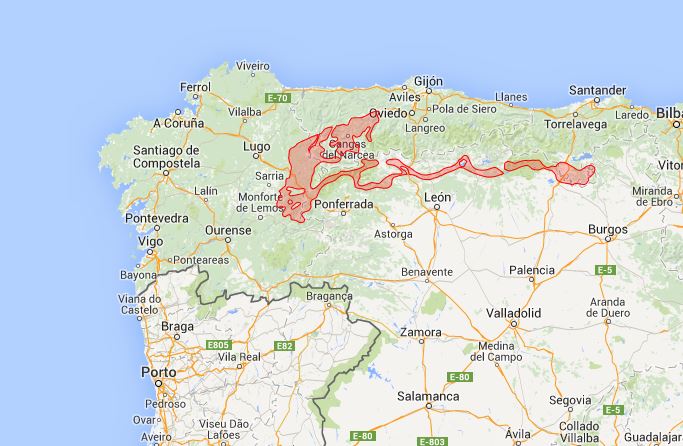
F24 Melampyro pratensis-Quercetum pyrenaicae Rivas-Martínez in Rivas-Martínez, T. E. Díaz, F. Prieto, Loidi et Penas 1984 (euscaldian); Linario triornithophorae-Quercetum pyrenaicae Rivas-Martínez, T. E. Díaz, F. Prieto, Loidi et Penas 1984 (orocantabrian).
|
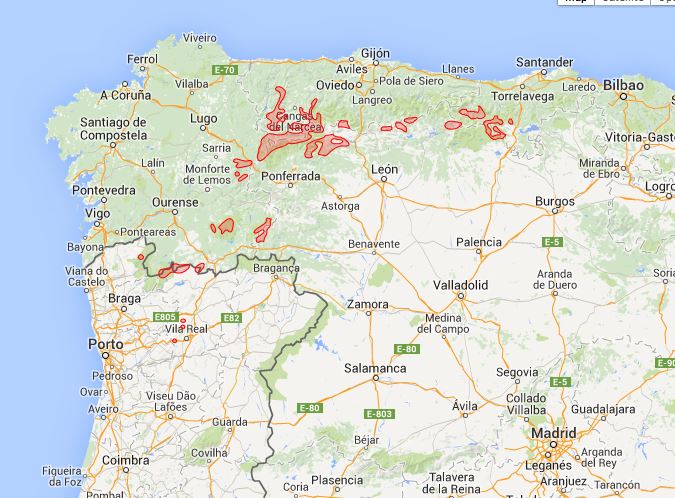
F25 Linario triornithophorae-Quercetum petraeae (Rivas-Martínez, Izco et Costa ex F. Navarro 1974) F. Prieto et Vázquez 1987; Luzulo henriquesii-Quercetum pyrenaicae (F. Prieto et Vázquez 1987) F. Prieto et Vázquez 1994.
|
The alliance Quercion pyrenaicae Rivas Goday ex Rivas-Martínez 1964 [1963] comprises the Quercus pyrenaica woodlands on acid soils, with their main area in the northern Iberian province. The formation subgroup G.4.1., with the units G64 to G70, belongs to this alliance. The Quercus canariensis woodlands of the formation subgroup G.4.4. on silicate rock are also close to this unit. As a submeridional unit, this alliance can be compared with the Castaneo-Quercion Soó 1964 (see below).
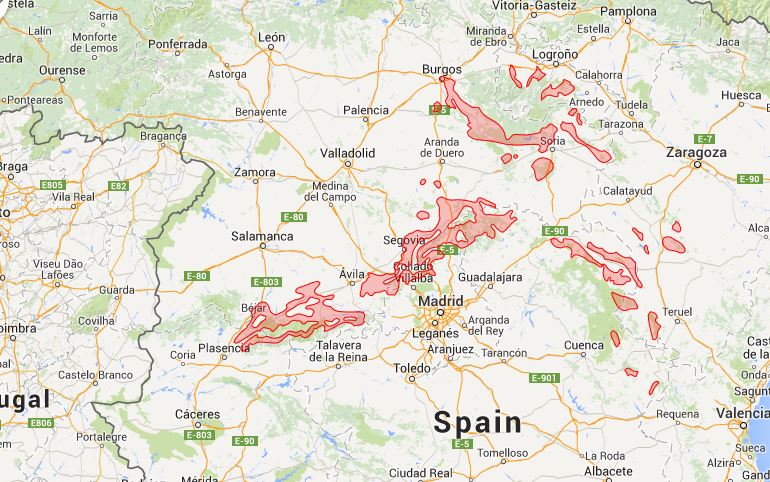
G64 - Central Iberian supra-Mediterranean Quercus pyrenaica-forests with Luzula forsteri
| 
G70 - Lusitanian-Extremadurian meso-Mediterranean Quercus pyrenaica-forests with Arbutus unedo
|
We
have decided to raise the former suballiances to the rank of alliances.
The different linkages to the vegetation zones and the oceanity of both
alliances should be pointed out: on the one hand temperate southern
Atlantic woodlands, mostly dominated by Q. robur; on the other hand submeridional northern Iberian woodlands, mostly dominated by Q. pyrenaica (see
map). In this way the situation is avoided whereby one alliance covers
two vegetation zones. Further clarification is still required as to
whether the western sub-Mediterranean Quercion pyrenaicae and the central to eastern sub-Mediterranean Castaneo-Quercion might be united within an order of sub-Mediterranean acidophilous oakwoods.
…
Macroclimatic factors (J. Cross)
The
distribution of this formation corresponds with climate type VI and the
more oceanic variant with type V-VI, as well as type V, based on WALTER
et al. 1975 (see Map 2). The large longitudinal and latitudinal range
of this formation indicates that it can tolerate remarkable variations
of climate. In the west, winters are mild with only a few frost days
(average of the coldest month in Great Britain and Ireland is
approximately 3-5 °C), summers are cool (average of the warmest month
approximately 16 °C or less) and the mean annual temperature lies at
approximately 10 °C. Cloud cover and relative humidity are high. This
pattern also applies in the north-west, oceanic areas of mainland
Europe. To the east the climate becomes increasingly more continental:
the precipitation decreases from more than 2000 mm on the west coasts of
Ireland and Britain, to 500-600 mm in eastern Poland and Ukraine and
there is a shift to a summer maximum. The winter temperatures decrease,
with averages of the coldest months ranging from 0 to -2 °C in north
Germany to -12 °C in Russia. In eastern areas continuous frosts and snow
cover can persist for a month or longer. The summers are warmer though
in the east with the averages of the warmest month lying between 16-20
°C and up to 23 °C in Russia. The annual mean temperature, however, is
lower than in the west, mean values being between 6.5 and 9 °C (and 5 °C
in the coldest region in north-east Poland). Southern Sweden has a
somewhat more continental climate than neighbouring areas in northern
Germany. Temperatures increase towards the south in the oceanic areas of
the formation. The highest values occur in south-western France
(average of the warmest month 20-22 °C, and the coldest month 4-6 °C);
on the Iberian Peninsula the summer temperatures are somewhat lower
(approximately 20 °C), but the winter temperatures are higher (6-10 °C).
The precipitation in south-west France shows a pronounced summer
minimum (lowest values around 500 mm). Precipitation increases again to
the west and exceeds 2000 mm locally in the mountains of north-west
Spain. The Insubrian distribution area shows similarities with the
north-west of the Iberian Peninsula with high annual precipitation
(1350-2000 mm), but the temperature of the warmest month, with 25 °C, is
much higher.
Site conditions (J. Cross)
Acidophilous
oak forests occur mainly on acidic, base-deficient soils (podzols,
gley-podzols, base-poor brown earths and ranker). The soils themselves
have developed as a rule from Palaeozoic and Mesozoic siliceous rocks
(sandstone, greywacke, slate, gneiss, other acid metamorphic rocks and
granite) or – in the Central and eastern European lowlands – from
Tertiary and Pleistocene unconsolidated material (moraines, outwash
plains), river sediments or – very locally – drained mires. The soils
are acidic to strongly acidic with a pH of < 4.5 to 5.5, oligotrophic
to oligo-mesotrophic, locally mesotrophic, very permeable, fresh to
dry, locally water-logged. In the uplands the soils can range from
shallow to rocky. In Central and southern Europe, thermophilous forms
occur on warm, sunny, south-facing slopes.
Role in structuring the landscape (J. Cross and U. Bohn)
On
the margins of, and outside the natural range of beech forests, as in
the north-west of the Iberian Peninsula, south-west France, Ireland and
Great Britain, acidophilous oak forests are prominent and characteristic
components of the siliceous uplands, as well as of the nutrient-poor
sandy lowlands (for example the ‘Landes’). In contrast, in western and
Central Europe, i.e. the domain of beech forests, they occur only on
sites inimical to the development of beech. These include highly
oligotrophic and dry sandy soils, water-logged sandy and silt soils of
the lowlands and hills, as well as shallow to rocky, summer-dry to
xerothermic sites within the siliceous uplands. In areas with extensive
stands, such as the Carpathian foothills in Romania, Bohemia,
Piedmont-Insubria and Croatia, they are restricted to the
colline-submontane altitudinal belts below the acidophilous beech
forests. At the margins of, and beyond the range of beech forests in the
central east European lowlands, the acidophilous oak forests occupy
oligotrophic dry to water-logged sandy locations. Here, the tree layer
regularly contains Scots pine and often also spruce. The natural
vegetation complexes include pine forests (on extremely sandy soils,
such as inland dunes), pine-bog forests, wooded raised bogs, birch and
alder carrs (on very wet, peaty sites) as well as oak-hornbeam and
small-leaved lime-pedunculate oak forests on more nutrient-rich sites.
In
addition to the above mentioned azonal forests, the following
vegetation types are also encountered within the formation: mixed
oak-ash forest on base-rich soils (e.g. F19), oak-alder-ash in river
flood plains, oligotrophic mires, raised bogs and (in the euatlantic
area) blanket bogs on sites with poorly drained peats. On naturally
rocky and dry sites, rock heaths, rock scrub and siliceous dry
grasslands occur as adjacent communities.
State of preservation, land use, substitute communities (J. Cross and U. Bohn)
Traditionally, acidophilous oak forests have been used for coppice, but small, near-natural stands
have
persisted in many regions, in Central Europe primarily on extreme
sites. Many of the seminatural coppice stands have been allowed to
develop into high forest, or they have been replaced by plantations of
Scots pine (Pinus sylvestris) or more rapidly growing, exotic conifers, such as Douglas fir (Pseudotsuga menziesii).
Extensive areas of the oligotrophic and poorly productive soils are
unsuitable for agriculture without the regular addition of organic or
mineral fertilisers. In the past, after thinning and forest clearance,
the areas were typically grazed, usually by pigs and sheep. As a
consequence large areas of these forests have been converted to dwarf-
shrub heaths, acidic grasslands, open siliceous grasslands, or –
depending on human activities – to dense stands of bracken. Recently,
many of these heaths have been afforested with conifers, although
grazing still plays an important role in certain areas. Nowadays, in
some areas former grazing practices are being reintroduced for nature
conservation purposes in nature reserves dedicated to preserving the
traditional, cultivated landscapes and their vegetation, often using old
breeds of domesticated animals Arable land is concentrated on dry areas
of the lowland units, where arable crops such as barley, oats, rye,
potatoes and sometimes corn are cultivated.
Nature conservation (J. Cross)
Acidophilous
oak forests remain mostly as small, often derilict stands of old
coppice or high forest and are currently highly endangered in many
areas. The principal threats are from, grazing, clear-felling,
afforestation with conifers and conversion into arable land. Air
pollution and eutrophication from fertiliser drift, especially nitrogen,
represent major problems, particularly in Central and northern Europe,
as they lead to the proliferation of nitrophilous species and brambles.
Overpopulation by game species and (particularly in Great Britain and
Ireland) overgrazing by sheep prevent the regeneration of these forests.
As a consequence, many stands lack a dwarf shrub layer and only possess
a severely degraded herb layer, while the tree layer is often
even-aged. In certain areas, fire also presents a risk, while
recreational use can also present a problem locally. In the wetter parts
of Great Britain and Ireland, the invasion of Rhododendron ponticum,
and locally also beech, pose a serious threat to these forests. Many
countries have established nature reserves, national parks or nature
parks for protecting valuable, semi-natural forest stands and substitute
communities worthy of protection (particularly dwarf shrub heaths,
mat-grass communities, fields of Corynephorus). However, these fall far short of preserving and restoring sufficiently large and representative stands of all forest types.
Classification into subunits (J. Pallas)
In
the overall legend the acidophilous oak forests are subclassified
mainly according to the altitudinal belts they occupy, which are
arranged basically from north to south:
– lowland-colline types (F1-F13)
– colline-submontane types (F14-F22)
– montane-altimontane types (F23-F26).
Within
these belts, the mapping units are, as a rule, arranged from west to
east, from markedly oceanic to subcontinental climates. This is manifest
in the corresponding floristic changes. In addition, ecological
variants are also identified, i.e. hygrophilous (F3, F9, F22), xerothermic (F17 p.p., F18, F20 p.p.) and thermophilous types (F21).
However, in Table 12 and the following text, the classification is based on on synchorological criteria, whereby the altitudinal belts are arranged on a regional basis. The sequence begins with the southern Atlantic units of northern Iberia (F14, F15, F23-F26), which most clearly demonstrate the thermal-oceanic character of this area. This is followed by the southern Atlantic mapping units of France (F4-F7), followed by the sub-Atlantic Burgundian unit F16 and the sub-Atlantic Rhenanian unit F18, with the submeridional Piedmontese-Insubrian unit F17 being placed alongside. The primarily northern temperate units are arranged from west to east, from the extremely hygric oceanic unit of Ireland (F1) to the Central European and Sarmatian units (F12, F13) that already show subcontinental traits. The last section of the table is formed by southern temperate Central European and pre-Carpathian (F20, F22), as well as the submeridional Illyrian (F21), units.
Table 12 is organised according to the vertical struture (tree, shrub, liane, herb and moss layers).
Within
the layers the general character species are listed first. These are
followed by the species characteristic for the Atlantic and sub-Atlantic
units as well as those characteristic for the Central
European/Sarmatian units, consistent with the main dividing line between
the oceanic and subcontinental units. Within each of these groupings
the species are arranged according to their zonal chorological linkages,
i.e. according to their affiliation to the major distribution type
groups. Differential species based on habitat and altitude are grouped
together into their own groups. The literature regarding the individual
mapping units that was studied for compiling Table 12 is cited in the data sheets.
Geologia, Geomorfologia e Solos
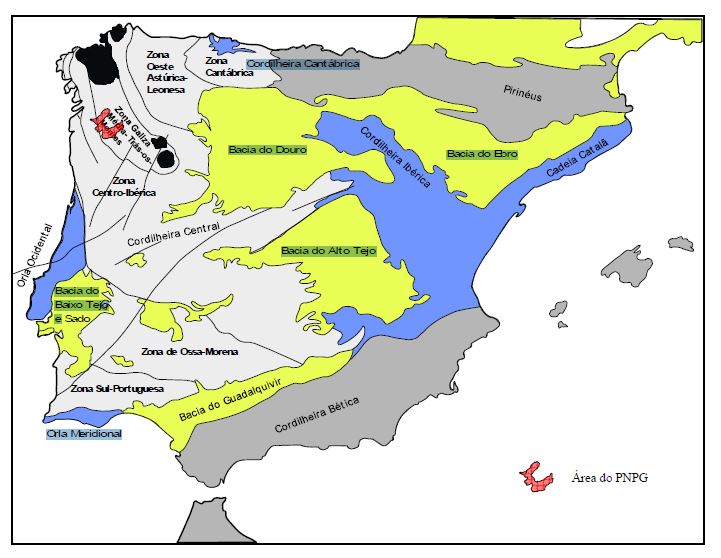
Enquadramento
geográfico do PNPG nas unidades morfoestruturais da Península Ibérica e
nas zonas definidas para o Maciço Ibérico (adaptado de Ribeiro et a!,
1979; Farias et a/., 1987)
Uma resumida caracterização geológica do Parque Nacional da Peneda-Gerês encontra-se numa página da Universidade do Minho sobre o Parque Nacional da Peneda-Gerês sobre dos Aspectos Geológicos e Geomorfológicos do PNPG:
A área do PNPG integra-se no Maciço Hespérico ou Maciço Ibérico que constitui uma das unidades estruturais da Península Ibérica e um segmento da Cordilheira Varisca da Europa. A edificação desta estrutura, pela actuação de forças compressivas, inicia-se no Devónico, há cerca de 380 Ma (milhões de anos), tendo-se prolongado até ao Pérmico (280 Ma) - orogenia Hercínica ou Varisca.
O Maciço Ibérico apresenta-se zonado, definindo-se habitualmente cinco zonas com características paleogeográficas, tectónicas, magmáticas e metamórficas distintas. A área do PNPG situa-se na Zona Centro-Ibérica (a zona mais interna da Cadeia Varisca). Esta zona é genericamente caracterizada pela existência de rochas muito deformadas e afectadas por elevado grau de metamorfismo e ainda pela predominância de rochas graníticas.
Uma descrição do património geomorfológico-glaciário e da geodiversidade do PN Peneda-Gerês encontra-se numa tese de mestrado de Luciana Peixoto de 2008 sobre: O património geomorfológico–glaciário do Parque Nacional da Peneda Gerês: proposta de estratégia de geoconservação.
Resume da tese
O Parque Nacional da Peneda–Gerês (PNPG) apresenta, além de muitas outras características que o tornam ímpar, uma elevada geodiversidade, sendo que, alguns dos aspectos apresentam valor patrimonial.
Desde o século XIX que é referida a possibilidade de terem existido processos glaciários nas serras do PNPG, tendo esta sido confirmada com os indícios encontrados na área do parque. As formas glaciárias são de escala variada, ocorrendo geoformas quer de erosão quer de acumulação, nomeadamente dos tipos: circo; vale glaciário; rochas aborregadas; estrias; polimentos e arrancamentos; moreias e blocos erráticos.
A inventariação dos Locais de Interesse Geomorfológico–Glaciário (LIGGs) resultou dum trabalho prévio de reconhecimento dos tipos de património geológico. Entre estes, os locais de maior valor patrimonial no PNPG são, de categoria, geomorfológicos do tipo glaciário.
A
relevância dos LIGGs foi obtida por quantificação quer no âmbito
nacional e internacional quer à escala regional e local. A quantificação
(Q) teve como critérios base o valor intrínseco (A), o seu uso
potencial (B) e a necessidade de protecção (C).
Dos LIGGs identificados apenas cumprem os requisitos para serem considerados de âmbito nacional e internacional o Circo glaciário de Cocões de Concelinho, o Vale glaciário, sector com perfil em U do alto Vez e os Complexos morénicos no vale da ribeira do Couce e próximos da Lagoa do Marinho. O Circo glaciário de Cocões de Concelinho encontra-se em primeira prioridade no que se refere à elaboração de uma estratégia de geoconservação, seguindo-se-lhe o Vale glaciário, do alto vale do Vez. O valor (Q) dos Complexos morénicos é inferior ao do Circo glaciário de Cocões de Concelinho mas são formas associadas a este e foram integradas na estratégia de geoconservação.
Dos geossítios Circo glaciário de Cocões de Concelinho e complexos morénicos foi elaborada uma visita virtual a disponibilizar na internet, uma vez que, a sensibilidade do local à presença do Homem é elevada.
No Alto vale do Vez, uma vez que existe um trilho bem marcado, desenvolveu-se um guião que complementa esse percurso pedestre.
Este trabalho pretendeu ser um contributo voluntário para que a geodiversidade seja cada vez mais valorizada pela sociedade actual e um ponto de partida para muitos outros estudos no tema da Geoconservação que na área possam ser desenvolvidos

Veja à seguir: 2.13.25 - Trás-os-montes e Alto Douro
[1] Moritz Willkomm (1852): Die Strand- und Steppengebiete der iberischen Halbinsel und deren Vegetation. Leipzig 1852. (Download)
[2] In: Leonel de Vieira. Portugal Património: Guia - Inventário (em ). Casais de Mem Martins, Rio de Mouro: Círculo de Leitores, 2007. Capítulo G02. p. 10. vol. II. ISBN 978-972-42-3908-8.
[5][http%3A%2F%2Fportal.icnb.pt%2FNR%2Frdonlyres%2F2CCF76F8-2288-4A03-B11E-1828DE1282C5%2F0%2FCARACTERIZACAOPATRIMONIO.pdf
Revisão do plano de ordenamento do parque nacional da Peneda-Gerês]
[6] À Descoberta do Parque Nacional Peneda-Gerês www.destinoseviagens.com. Visitado em 13 de Janeiro de 2010.
[9] DOWNLOAD - EuroVegMap - CD Interactico da Vegetação Natural Potencial da Europa - a instalação oficial deu erro em Windows 8.1
[11] For a definition of Middle Europe: see chapter 2.3, section ‘Floristic regions of Europe’ and chapter 5.4.1.