
“Flowers of South-West Europe - a field guide” - de Oleg Polunin e B.E. Smythies
“Revisitas” de regiões esquecidas no tempo - “Plant Hunting Regions” - a partir de uma obra de grande valor para o especialista e amador de botânica como da Natureza em geral.
Por
Horst Engels, Cecilia Sousa, Luísa Diniz, Nicole Engels, José Saraiva, Victor Rito
da
Associação “Trilhos d’Esplendor”
2.13 The Northern Serras of Portugal
2.13 As Serras do Norte de Portugal
2.13.2 Serra da Estrela
|
Folha de Cálculo: Flora da Serra da Estrela
(Lista provisória de plantas vasculares e não-vasculares)
|
Bases de Dados:
|
Mapas das Serras do Norte de Portugal:
|
2.13.2 Serra da Estrela
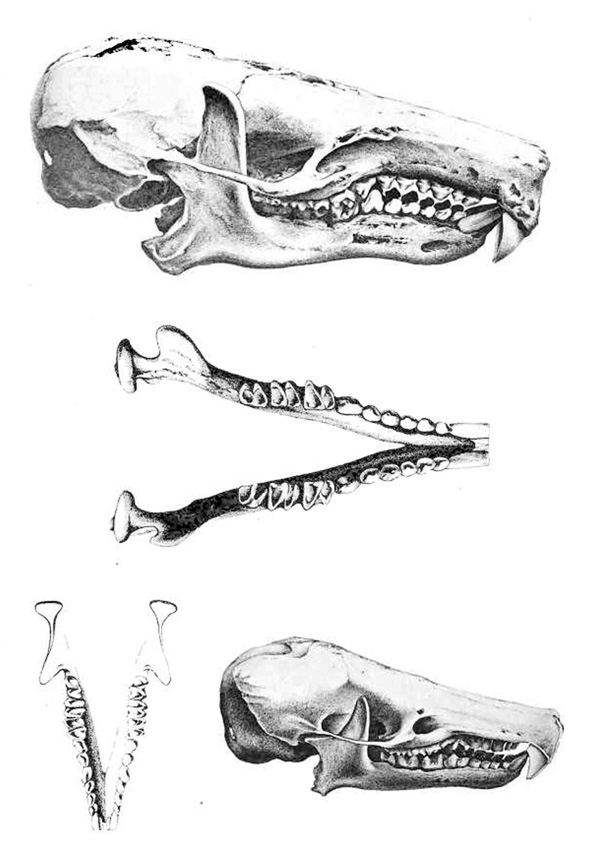
Galemys pyrenaicus - Crânio
2.13.2d4 - Aspectos glaciários - Filogeografia (Galemys pyrenaicus)[1]

Topeira da Água (Galemys pyrenaicus)[2]
No seguinte vamos comparar resultados de um estudo morfológico (2006)[3] sobre a variabilidade craniométrica de Galemys pyrenaicus com resultados da análise genética (filogeográfica) sobre esta espécie (2013)[4]. Será interessante a observação da resolução filogeográfica obtida nos dois métodos e ver até que ponto os resultados genéticos e morfológicos conduziram às mesmas ou à conclusões diferentes sobre a filogeografia e história biogeográfica desta espécie.
Para quem está interessado nas técnicas aplicadas nestes dois estudos, aqui duas referências de livros. Para os estudos da estatística multivariada tem interesse o livro: “Multivariate Analysis of Ecological Data using CANOCO” de Jan Leps e Petr Smilauer (2003) e para os estudos da filogeografia um livro que foca bem a genética de conservação de espécies: “Population Genetics for Animal Conservation” (2009).
A) O estudo morfológico …
No estudo morfológico sobre Galemys pyrenaicus que habita o norte-oeste da Península Ibérica inclusivé os Pirenéus foram usados 209 crânios de diversas coleções museais. Foram tirados 14 medidas craniais e dentais nos exemplares.
Miller (1914) distingue 2 subespécies, Galemys pyrenaicus subsp. pyrenaicus e G. pyrenaicus subsp. rufulus através do tamanho do corpo e comprimento condylo-basal (medida para o tamanho do crânio) e através de uma diferença na cor da pele (G. pyrenaicus subsp. rufulus, além de ser maior no tamanho, mostra segundo Miller uma cor ligeiramente mais acastanhada do que G. pyrenaicus subsp. pyrenaicus).
No entanto, estas diferenças não podiam sempre ser confirmadas nos estudos posteriores sobre esta espécie. Alguns autores não conseguiram confirmar as diferenças na cor, outros não detectaram diferenças nenhumas ou apenas ligeiras no tamanho.
O estudo craniométrico recente que utiliza além das técnicas estatísticas habituais como análise de variância também métodos de estatística multivariada como cluster-análise (UPGMA) e métodos de ordenação, multiple group principal component analysis (MGPCA) e análise canonica de variadas (CVA).
Vemos aqui os resultados da análise de cluster UPGMA.
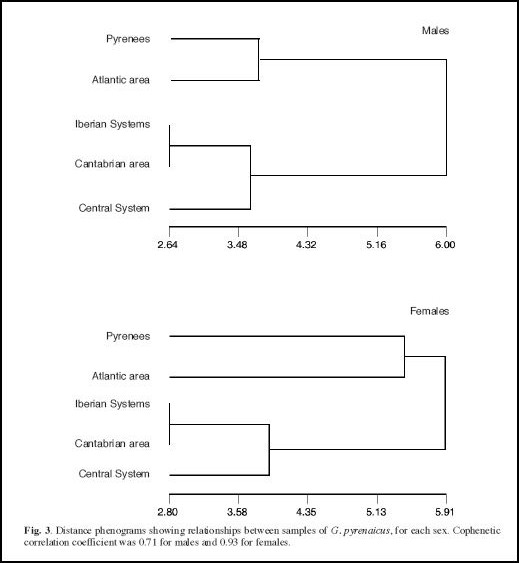
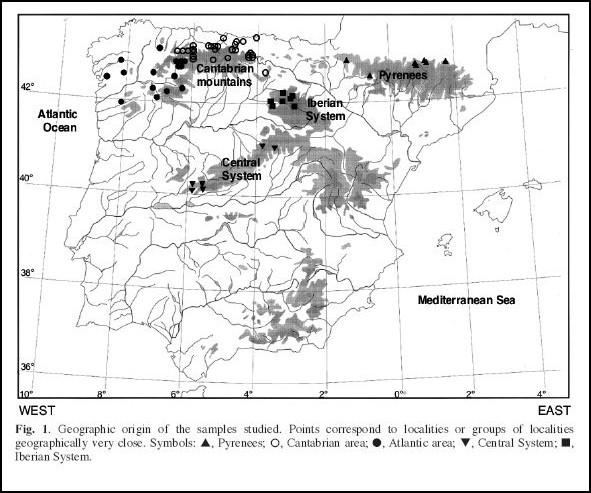
O resultado desta análise cluster demonstra um agrupamento de populações dos Pirenéus com populações da “área atlântica” - versus populações do Sistema Ibérico, da “área cantábrica” e do Sistema Central em ambos os sexos. Os resultados desta análise de cluster e das análises de variância parecem confirmar pelo menos a existência de 2 grupos maiores nesta espécie cuja distribuição correspondente parcialmente à distribuição das duas subespécies anteriormente descritas. No entanto, o agrupamento de exemplares da “área atlântica” com exemplares do Pirenéus diverge da diagnose dos autores anteriores.
Por outro lado, análises de estatística multivariada de ordenação como análise dos componentes principais ((MG)PCA) e a Análise Canónica de Variadas (CVA) permitem extrair e separar um factor “de tamanho” (o primeiro factor) que domina normalmente as análise estatísticas morfológicas dos restantes factores. Os restantes factores representam assim diferenças nas proporções e são normalmente interpretadas como forte indicadores de diferenças genéticas. O resultado das análises de MGPCA e CVA deste estudo revela 4 factores significativos, o primeiro fortemente (negativamente) correlacionado com “tamanho (size)”, os restantes 3 factores como factores de “proporções (form)”
Assim o resultado da análise morfológica nos Galemys pyrenaicus esta en conformidade com os estudos de Miller e de outros autores posteriores que confirman diferenças de tamanho entre as duas subespécies. Além disso as técnicas de MGPCA e CVA revelaram também diferenças nas proporções entre as subespécies. No entanto, o agrupamento das populações dos Pirenéus com aquelas da “área atlántica” (em relação ao tamanho pequeno) vem de surpresa.
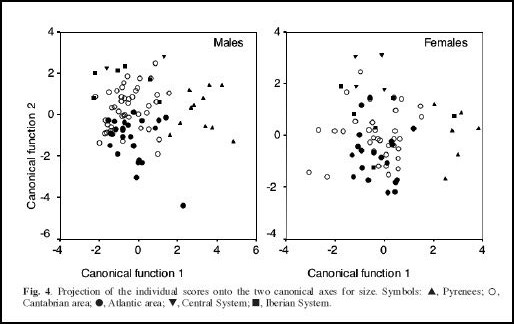
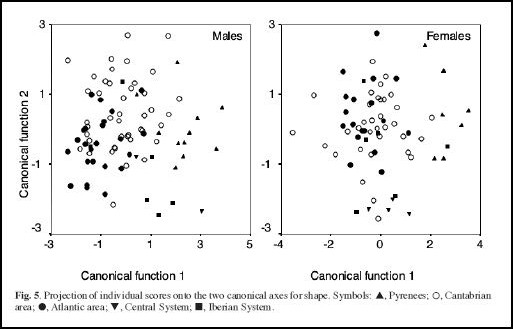
À primeira vista, o agrupamento das populações dos Pirenéus com as populações da “área atlántica” surgere que as populações destas duas áreas geográficas deviam pertencer a mesma subespécie pyrenaica, mas os resultados das análises de ordenação (que permitem excluir um factor “tamanho” dos restantes factores de “proporções”) indicam que apesar do tamanho menor e parecido nas populações dos Pirenéus e da “área atlántica”, haver diferenças significativas nas proporções entre as amostragens destas duas regiões.
Assim, o resultado morfológico parece suportar pelo menos a diagnose inicial da existência das duas subespécies e aproximadamente da sua distribuição geográfica suposta:
“We conclude from these arguments that there is an overall morphological skull differentiation between the subspecies of the Iberian desman described in the literature. Differences observed between the Pyrenean population of Galemys and those from other Iberian regions could be explained by their geographical isolation, caused by changes that occurred during the last glacial event (Würm). Specifically, climate and vegetation changes in that period resulted in two, well-differentiated faunal groups, observed from 24000 to 12000 BP (B a r y s h n i k o v & M a r k o v a 1992): “Mammoth tundra-steppe assemblage”, and “Mediterranean mountain open woodlands assemblage”. A similar process may have occurred in other mammal species with Pyrenean populations taxonomically different from those in the rest of the Iberian Peninsula, such as Microtus arvalis meridianus, Chionomys nivalis aquitanius, Clethrionomys glareolus vasconiae, Arvicola terrestris monticola, or Capra pyrenaica pyrenaica (see e.g. P a l o m o & G i s b e r t 2002). It is clear that in the case of the Iberian desman results from other sources may help to establish the taxonomic identity of the two subspecies classically recognised for this insectivore..
O estatuto taxonómico e a situação filogeográfica de Galemys pyrenaicus das “áreas atlánticas e cantábricas” da Península Ibérica fica assim pouco conclusivo neste estudo craniométrico.
Extratos de textos:
Introduction
The Iberian desman, Galemys pyrenaicus, is a member of the Talpidae family and is endemic to the mountain areas of the northern half of the Iberian Peninsula. The species was first described by E. G e o f f r o y S a i n t - H i l a i r e (1811), who assigned a specimen from the northern side of the Pyrenees to Mygale pyrenaica. The generic name Galemys was used for the first time by K a u p (1829), though it was not generally used until M i l l e r ’ s 1912 Catalogue. Meanwhile, G r a e l l s (1897) attributed desmans from the Sierra de Guadarrama (central Spain) to a new taxon, Myogalea rufula, on the basis of its more yellow shading on its back, legs and nails than the Pyrenean type specimen. M i l l e r (1912) recognised one species of the Iberian desman, Galemys pyrenaicus, with two subspecies, pyrenaicus and rufulus. He maintained that it occurred in the Pyrenees and adjacent parts of southern France, and probably also in north-eastern Spain north of the Ebro river; its presence in Asturias was also suggested. According to M i l l e r (1912), the form rufulus was distributed in central Spain, south of the Ebro Valley. To distinguish the two subspecies, this author indicated the larger hindfoot, larger condylobasal length, more massive skull, and slightly lighter fur colour of rufulus. This taxonomic pattern was followed by C a b r e r a (1914), who mentioned that rufulus was larger than pyrenaicus.
The characters distinguishing pyrenaicus and rufulus indicated by C a b r e r a (1914) were also the hindfoot and condylobasal lengths. Nevertheless, he found no differences in fur colour
between the two forms.
N i e t h a m m e r (1970) indicated that the body and skull sizes of the specimens from Burguete (western Pyrenees) were, in general terms, slightly smaller that those from other zones of northern (Sierra de Cameros in the Iberian System, and Cantabria) and central Spain (Guadarrama). He specified that these differences were most relevant in the condylobasal length and that the mean body weight recorded in his rufulus specimens was clearly higher than the values mentioned by R i c h a r d & V a l l e t t e V i a l l a r d (1969) for animals from the French Pyrenees. Nevertheless, N i e t h a m m e r (1970), like C a b r e r a (1914), found no differences in the fur colour of pyrenaicus and rufulus. Using a full sample of specimens from all geographic zones, he indicated that both sexes were very similar in their body and skull dimensions as well as in fur colour.
P a l m e i r i m & H o f f m a n n (1983) mentioned the two subspecies, although they pointed out that they had few morphological differences. They based themselves on N i e t h a m m e r ’ s (1970) observations to state there was no size dimorphism between sexes.
In J u c k w e r ’ s (1990) review of the species, the validity of pyrenaicus and rufulus was maintained on the basis of the size characteristics mentioned by M i l l e r (1912), C a b r e r a (1914), and N i e t h a m m e r (1970). J u c k w e r (1990) found no significant differences in body or skull size between males and females from the Sierra de Cameros. In a more detailed morphological analysis of these forms, G o n z á l e z - E s t e b a n et al. (1999) concluded that the identity of the two subspecies was doubtful since body and skull sizes of desmans from the western Pyrenees fell completely within the range observed in specimens from more western zones (Iberian System mountains, Cantabria, Asturias, León, and Galicia).
Pooling specimens from different geographic zones in a common sample, these authors found no significant differences in body and skull sizes between sexes, corroborating N i e t h a m m e r’ s (1970) and J u c k w e r ’ s (1990) findings. According to G o n z á l e z - E s t e b a n et al. (1999), fur colour varies geographically in such a way that specimens from Pyrenees, Basque Country, and Iberian System are noticeably darker than those from most western sites (Asturias, León, and Galicia). Nevertheless, they specified that these morphological patterns did not correspond to the classic differentiation between the subspecies pyrenaicus and rufulus.
We have been compiling craniometric data on the Iberian desman for several years.
Preliminary analyses of these data suggested that several biometric relationships between geographic samples and sexes differed from certain results reported in the literature.
Additionally, no information on skull shape variation in the distribution area of the species has ever been reported. With all this in mind, the aims of this paper are: i) to analyse the extent of sexual dimorphism in skull size of the Iberian desman at a local level; ii) to review geographic variation in skull size; and iii) to determine for the first time the intraspecific relationships in skull shape of G. pyrenaicus in most of its distribution area. It is worth
remembering that the Iberian desman is included in Appendices II and IV of the Habitats and Species Directive (see B l a n c o & G o n z á l e z 1992) and has recently been listed as Endangered in the Atlas of the terrestrial Mammals of Spain (N o r e s et al. 2002).
Therefore, information on the morphologic characteristics of the species throughout its geographic range could be of interest to conservation and management programmes aiming to preserve the full genetic variability of this endangered insectivore.
|
Discussion
Age and sex variation
The lack of sexual dimorphism in body and skull sizes of the Iberian desman has been reported in several studies (N i e t h a m m e r 1970, J u c k w e r 1990, G o n z á l e z - E s t e b a n et al. 1999). Nevertheless, it should be emphasised that this assessment was based on the analysis of animals coming from a specific location (Sierra de Cameros; J u c k w e r 1990) or on general observations based on pooling of specimens from different geographic zones (N i e t h a m m e r 1970, G o n z á l e z - E s t e b a n et al. 1999). Our results showed that, in all geographic samples, skull size is in general slightly larger in males than in females. Statistical analyses revealed that, although in all samples mean values of many parameters did not differ significantly between sexes, sexual monomorphism in skull size seems not to be the
rule in the species. In fact, the degree of differentiation between males and females depended on the geographic sample analysed. Thus, although males and females from the Pyrenees were very similar in all cranial variables, several significant differences between sexes were recorded in desmans from other areas. Specifically, in the Cantabrian area sample differences occurred in 43% of the parameters considered; as for the other variables, mean values were always higher (but not significant) in males. In the remaining samples (Iberian System, Central System, and Atlantic area), differences attained statistical significance only in a few characters.
Available data concerning the biology of the species do not allow us to explain the causes of the slight sexual dimorphism in skull size or of the different patterns we found throughout its distribution area. Information on the social organization, spatial distribution, and activity patterns of the species (S t o n e 1985,1987a,b) indicated that paired adults defend their shared territory from neighbouring desmans by patrolling, surveillance, and scent-marking, and that in such pairs males cover more territory than females. Likewise, the home ranges of neighbouring males overlap to a small extent, but the territory of a male and that of a neighbouring female do not overlap (S t o n e 1987a). Paired males appear to
invest more time defending the borders of their shared home range than do females, which spend most of their time within the centre of the range. Though females also participate in territorial defence, their activities are concentrated in this central region. Despite some
degree of territorial overlap between neighbouring desmans, their mutual avoidance, achieved through a system of temporal range utilization, means that there are few agonistic encounters (S t o n e 1987a,b). Therefore, the slight sexual dimorphism in skull size detected could be related, at least in part, to these patterns of social ecology.
As for the geographic differences in the extent of dimorphism here reported, we tentatively suggest that the effect of different sample sizes underlies these results. Thus, whereas most significant differences and/or a higher degree of sexual differentiation occurred in the larger samples from the Cantabrian and Atlantic areas, differences between males and females were mostly undetectable in the other samples. In consequence, we would expect that, if more specimens were added to these latter samples, sexual differences would attain statistical significance in more skull variables. Therefore, we conclude that in the Iberian desman the skull is on average slightly larger in males than in females, but these differences attain statistical significance only when a larger sample size of a specific population is examined. This is probably the reason why sexual dimorphism in skull size has been ruled out in analyses based on small size samples or in multivariate analyses based on a whole sample composed of subsamples with heterogeneous composition of sexes (e.g. N i e t h a m m e r 1970, J u c k w e r 1990, G o n z á l e z - E s t e b a n at el. 1999). To test the latter question, we performed an additional multivariate analysis of variance (MANOVA ) on our whole sample, taking age, sex, and geographic origin as factors. Unlike the results here obtained by univariate analyses, MANOVA did not show overall significant difference in skull size between sexes (Hotelling’s T2 = 0.201; F = 1.782; p = 0.121). In the light of our results, we recommend treating males and females separately when comparative analyses of skull morphology between geographic samples are undertaken.
We found no significant differences in skull dimensions between the four age classes established in G. pyrenaicus. This is consistent with the results by G o n z á l e z - E s t e b a n et al. (1999) and supports the idea that, as occurs in other Erinaceomorpha species (see F o r e s m a n 1994), when young animals leave the nest they have already reached adult skull size.
Geographic variation
Our one-way ANOVA , after the Bonferroni adjustment, revealed that the skull size of G. pyrenaicus varies significantly over its distribution area. In general terms, desmans (both males and females) from the Pyrenees were smallest and those from the Cantabrian area, Iberian System, and Central System were largest. Likewise, canonical analyses showed an important distinction in both size and shape between the Pyrenean animals and the specimens traditionally assigned to rufulus. Specifically, excluding factor size, Pyrenean animals are characterised by their robust interorbital region and, additionally, males show a wider rostrum and females a shorter coronoid process. Although slightly larger, animals from the Atlantic
area were very similar in size to the specimens from the Pyrenees. The relatively small size of the desmans from the Atlantic area was also reported by G o n z á l e z - E s t e b a n et al. (1999). Specifically, these authors found that animals from the western Pyrenees had an average size that was intermediate between desmans from westernmost Spain (Galicia and western Leon) and those from the Cantabrian region (Cantabria, Asturias, and northern Leon) and the Iberian System. Based on this and on their findings on geographic variability in the fur colour of the species, these authors argued against a subspecies differentiation founded on body and skull sizes. Thus, G o n z á l e z - E s t e b a n et al. (1999) suggested that the morphological variability of the species is related to several local factors, such as low population densities, the territorial behaviour of the species, and the partial or total isolation of populations. Although the effect of these factors at a local level cannot be discarded, our results do not discredit so radically the existence of morphometric differentiation between pyrenaicus and rufulus. Thus, excepting the animals from the Atlantic area, desmans from the Pyrenees are clearly smaller in size than those from the other areas considered here. In fact, the specimens assigned to rufulus lead one to think that a geographic cline in skull size exists as follows: smaller desmans in the west (Galicia), larger ones in the mountains of central Spain (Central System), and intermediate ones in the Cantabrian region and Iberian System. Specifically, animals from Galicia were significantly smaller in many skull variables than those from the other samples attributed to rufulus. Although these results and the relatively high overlap in skull dimensions between Pyrenean and Galician animals seem at first to contradict the taxonomic identity of pyrenaicus and rufulus, our analyses of skull size and shape by MGPCA and CVA revealed a substantial distinction between these forms, both in males and in females. In particular, in the corresponding canonical plots for size and shape, component scores of males and females from the Atlantic area fall within the range of variation of the other samples of rufulus, and appeared separate to a large degree from Pyrenean desmans. It is worth mentioning that the relatively low mean values recorded in the sample from the Atlantic area were mainly due to the very small skull size of the specimens coming from a specific locality, Saa de Incio (for example, CB L: males, mean = 33.50, sd = 1.07, range = 32.3-34.5, n = 5; females, mean = 32.45, sd = 0.83, range = 31.4-33.3, n = 4. ML: males, mean = 21.26, sd = 0.73, range = 20.3-22.0, n = 5; females, mean = 20.67, sd = 0.54, range = 19.9-21.1, n = 4). Therefore, the similarity in size of certain skull measurements between Pyrenean and some Galician desmans may be due to the action of local factors. We conclude from these arguments that there is an overall morphological skull differentiation between the subspecies of the Iberian desman described in the literature. Differences observed between the Pyrenean population of Galemys and those from other Iberian regions could be explained by their geographical isolation, caused by changes that occurred during the last glacial event (Würm). Specifically, climate and vegetation changes in that period resulted in two, well-differentiated faunal groups, observed from 24000 to 12000 BP (B a r y s h n i k o v & M a r k o v a 1992): “Mammoth tundra-steppe assemblage”, and “Mediterranean mountain open woodlands assemblage”. In the Iberian Peninsula, the border between the two ran along the base of the southern slopes of the Pyrenees. Pleistocene remains of G. pyrenaicus are scarce, but they indicate the occurrence of a Pyrenean population in Würm times (A r r i b a s 2004) living in cold tundra, in contrast to the other Iberian populations, which lived in temperate Mediterranean scrub. A similar process may have occurred in other mammal species with Pyrenean populations taxonomically different from those in the rest of the Iberian Peninsula, such as Microtus arvalis meridianus, Chionomys nivalis aquitanius, Clethrionomys glareolus vasconiae, Arvicola terrestris monticola, or Capra pyrenaica pyrenaica (see e.g. P a l o m o & G i s b e r t 2002). It is clear that in the case of the Iberian desman results from other sources may help to establish the taxonomic identity of the two subspecies classicaly recognised for this insectivore.
|
….
B) O estudo genético …
No estudo genético de Galemys pyrenaicus fora usados 69 amostras fecais, amostras de teçidos de 63 exemplares de coleções de museus e 2 amostras de fragmentos osseos de coleções históricas. Assim foram usados no total 134 amostras de exemplares diferentes de Galemys pyrenaicus do norte-oeste da Península Ibérica (Tabela S1)[5].
Extrato da Tabela S1.
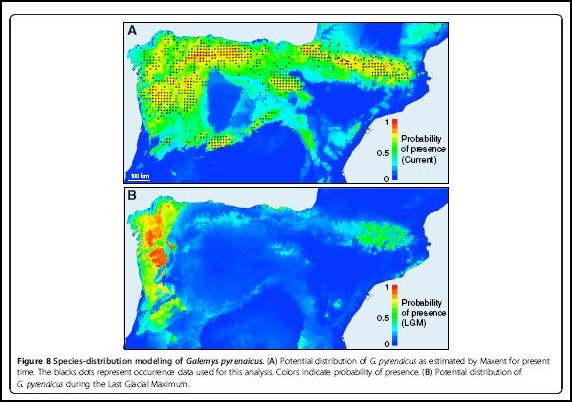
Species-distribution modeling of Galemys pyrenaicus. (A) Potential distribution of G. pyrenaicus as estimated by Maxent for present time. The blacks dots represent occurrence data used for this analysis. Colors indicate probability of presence. (B) Potential distribution of G. pyrenaicus during the Last Glacial Maximum.
O resultado da análise mitochondrial e agrupamento das 134 amostragens mostra 4 agrupamentos de populações de Galemys pyrenaicus.

Figure 1 Phylogeographic analysis of the mitochondrial sequences of Galemys pyrenaicus. (A) Map of the northern part of the Iberian Peninsula showing the 134 samples of G. pyrenaicus used in this study. The grayed area represents the historical species distribution according to different sources. Names of mountain ranges mentioned in the text are shown. Each sample is represented by a circle, but a few samples with the same coordinates cannot be discerned. Colors of the samples indicate the four different mitochondrial lineages recovered in the phylogenetic analyses (A1, A2, B1 and B2). The single locality with two samples belonging to two different lineages (B1 and B2) is shown with both corresponding colors. The purple line indicates the genetic barrier identified by the Monmonier’s Maximum Difference algorithm (the intersection with the species distribution area is shown with stronger color). (B) Haplotype genealogy of the concatenated mitochondrial sequences based on a maximum-likelihood tree. Circles represent haplotypes, with size being proportional to the number of individuals, and black dots representing intermediate, unsampled haplotypes. (C) Bayesian tree of the same sequences. Posterior probabilities for relevant clades are shown. The scale bar represents 0.001 substitutions/position.
Estes 4 grupos (linhas genealógicas) detectadas podem ser interpretadas como 4 linhas genealogicas distintas em que 2 linhas de cada são mais aparentadas. A distribuição geográfica dos 4 grupos sugere também uma linha de separação (barreira) genética entre populações cantábricas (B1) e dos Pirinéus (B2) e da “área atlántica” (A1) como entre populações de B1 e A2 no Sistema Norte Ibérico.

Figure 9 Schematic representation of the evolutionary history of Galemys pyrenaicus. The grayed area represents the historical species distribution. Sampled areas of the four mitochondrial lineages are shown in different colors. Hypothetical positions of glacial refugia are illustrated with dotted circles within the current distribution of each mitochondrial lineage. The size of the circles represents the relative importance of the refugia. Arrows indicate colonization routes from these refugia.
Tomando em conta toda informação obtida neste estudo genético consegue-se reconstruir através dos dados um scenário de história evolutiva de Galemys pyrenaicus com 4 grupos de isolamento genético durante um tempo de refúgio glacial com sucessiva expansão (colonização) a partir destes 4 centros refugais. Os centros A1 (Galiza) e A2 (Sistema Central) encontram durante a expansão pós-refugial de Galemys pyrenaicus os 2 distintos ramos de expansão do centro B1 (Sistema Cantábrico) que conduz à 2 barreiras de expansão (incluida como linha lilaz no primeiro gráfico), uma na área cantábrica e outra na área do Sistema Ibérico. O centro de expansão B2 (Baixos Pirinéus) encontra por seu lado o centro de refúgio B1 no caminho ao longo da costa atlântica na direcção da área cantábrica.
Esta informação suporta de facto a existência de 2 centros iniciais maiores (A1, A2 e B1, B2) de distribuição geográfica na Península Ibérica que deve ter conduzido ao processo de formação da duas subespécies distintas de Galemys pyrenaicus descritas morfologicamente:
For the moment, according to the genetic results and the corresponding type localities of the subspecies [20], the populations of mitochondrial group A would correspond to subspecies G. p. rufulus, and those of group B to subspecies G. p. pyrenaicus.
No entanto, a história evolutiva ainda é mais complexa: formaram-se 4 linhas filogenéticas distintas à partir de 4 supostos centros refugiais de Galemys pyrenaicus na Península Ibérica, possivelmente já antes da a última glaciação Würm.
Background
The genetic diversity patterns of species are a consequence of their evolutionary history (e.g. the existence of past refugia or vicariant geological events) and of contemporary constraints to dispersal (e.g. habitat fragmentation).
These processes are expected to give rise to specific phylogeographic patterns [1-3], the detection of which can be useful to infer the relative importance of different evolutionary and ecological forces. Pleistocene glaciations have been among the major drivers in shaping the genetic structure of species, particularly in the Northern Hemisphere [4-7]. The isolation of populations in separate glacial refugia generated, first, a subdivision of the genetic pool of species into clearly distinct lineages. Moreover, subsequent colonization of new areas caused a particular pattern of genetic diversity in which past refugia retained maximum levels of genetic diversity whereas recently colonized regions became more homogeneous [4]. However, current barriers to gene flow may be more determinant in the genetic structure of species inhabiting naturally fragmented habitats [8] or in species that have very specific ecological requirements, such as aquatic organisms [1].
The Pyrenean desman (Galemys pyrenaicus) is a small semi-aquatic mammal endemic to the northern half of the Iberian Peninsula. It occupies streams of clean and cold flowing waters with shallow but permanent water levels throughout the year, an habitat generally found in mountain areas. Its distribution is highly dependent on the presence of larvae of benthonic macroinvertebrates that the desman captures underwater. Adaptations to the aquatic life include a highly-mobile protracted snout, large hindfeet and a long tail with stiff hairs [9,10]. Like
many other specialists, the Pyrenean desman is an endangered species. For causes not clearly understood, it is undergoing significant declines across its whole geographic range. The situation has worsened during the last few years, particularly in the most southern populations, which have more Mediterranean climate. The decline of some populations has created a very fragmented distribution in this species [11]. The Pyrenean desman is legally protected in the four countries where it is present (Spain, Portugal, France and Andorra) and currently appears as “Vulnerable” in the IUCN Red List [12].
The Pyrenean desman forms part of the family Talpidae, which is included in the mammalian order Eulipotyphla (traditionally called Insectivora). Within Talpidae, the Pyrenean desman is placed within the subfamily Desmaninae together with the Russian desman (Desmana moschata), and therefore they are the only two extant representatives of this group of semi-aquatic mammals. Fossil data indicate that desmanines were much more diverse in the past [13,14] and the oldest fossil record is dated at 8.2 Myr [15]. The monophyly of desmanines is strongly supported by molecular data [16] and the divergence between both extant species has been estimated at around 10 million years (Myr) ago [17]. Therefore these two desman species are the last representatives of a unique lineage of specialist mammals that have experienced elevated extinction rates in the last few million years.
The Pyrenean desman is therefore an endemic, highly specialized, and relict species of great evolutionary and ecological interest. However, the genetic structure of the Pyrenean desman is yet to be investigated. Being a species with stringent ecological requirements, in which not all apparently favorable rivers are occupied [18], it is possible to hypothesize that the distribution of suitable habitats, very fragmented by their own nature, played a major role in structuring the genetic diversity of the species. For example, the genetic diversity could be partitioned, as in other organisms with strong aquatic requirements, according to major rivers or basins [1]. On the other hand, G. pyrenaicus is a polymorphic species in which two subspecies, pyrenaicus and rufulus, have been described according to differences in coloration and size. The validity of these subspecies and their distribution are still a matter of debate [19-21] but it has been postulated that these differences arose from geographic isolation during the Pleistocene glaciations [21]. Thus, the Pyrenean desman is a species of great interest on its own but it is also an ideal model to study how different ecological and evolutionary forces may have operated to establish the current distribution and genetic structure of species with strong ecological requirements.
To carry out a thorough genetic study of the Pyrenean desman, we first set up a noninvasive method of DNA extraction using droppings deposited on exposed rocks of the rivers it inhabits. Feces have previously been used to detect the presence of this elusive species [18,22] and represent a very valuable source of samples for genetic analyses across its whole distribution range. We favored a homogeneous sampling strategy in which samples were collected from as many localities as possible, rather than from discrete populations, to reduce biases in the delimitation of clusters and to better discern genetic diversity gradients [23]. Apart from feces, we also used tissue samples obtained from different biological collections as well as museum specimens. To assess the genetic diversity and the degree of connectivity between populations we used mitochondrial markers and nuclear introns. We show here how intron markers previously developed to be variable between closely related species and populations
[24] can provide crucial information to study the evolutionary history of species. Our results allowed us to obtain, for the first time, important insights about the population history of the Pyrenean desman, and may have critical implication for the conservation of this endangered species.
|
Discussion
Dating analysis of the mitochondrial lineages
We have been able to gather a number of solid pieces of evidence that show that the evolutionary history of G. pyrenaicus and the genetic structure of its populations were strongly influenced by the Pleistocene glaciations. Remarkably, the phylogeny of the mitochondrial genes exhibits a pronounced geographic pattern, with the presence of four lineages (grouped into two main phylogroups) that have a marked parapatric distribution. However, this structure is not by itself proof of the effects of glaciations as it may predate the Pleistocene. Therefore it is important to obtain an accurate dating of the splits of these lineages. Our dating approach allowed us to estimate that the split of the two most divergent mitochondrial phylogroups occurred 0.32 Myr ago and that the subsequent divergence of the two pairs of lineages concomitantly took place at around 0.23 Myr ago. Since sequence coalescence must be older than the population split, these dates represent the upper limit at which the desman populations started to diverge. Therefore, given these Middle Pleistocene lineage split times, it is very likely that the four desman populations evolved in four isolated glacial refugia, supporting the importance of the Pleistocene glaciations in the population structure of this species. Most probably, the populations started to diverge during earlier phases of the glacial periods and not necessarily in the last glaciation, explaining the deep mitochondrial divergences observed [59,60].
Since we did not have reliable fossils in desmanines, we had to use more external calibrations of mammals for our dating analysis. This analysis benefited from the nuclear introns, which allowed us to reconstruct a Bayesian tree of mammals calibrated with multiple fossils and to estimate the divergence date of Galemys and Desmana. The obtained date at 13.9 Myr was quite adequate to calibrate, in a subsequent step, the mitochondrial gene tree. On the one hand, this date is not as old as to present problems of saturation. On the other hand, it is not as recent as to suffer from the problems of coalescence, which can be exacerbated when dating very recent nodes of a gene tree (< 10 Myr) [61]. This calibration date was then introduced into a phylogenetic tree of the cytochrome b of talpids and, from this calibrated tree, we estimated the divergence time of the main Galemys mitochondrial lineages at 0.32 Myr. The evolutionary rate resulting with this approach for cytochrome b was 0.0224 substitutions/position/Myr. Although this evolutionary rate is line with those obtained for other mammalian groups [62,63], different dating approaches have led to much higher rates [64-66]. Also, actual quantification of mutation accumulation from pedigree data has shown more elevated evolutionary rates in mitochondrial genes, at least in humans [67]. It has been suggested that the possible existence of mutational hotspots and other problems [64] may cause that evolutionary rates can only be properly estimated in recent branches of a phylogeny. However, the extent of this effect is contentious [68,69]. Actually, it has been shown more recently that, in fact, lack of consideration of coalescence of ancestral polymorphisms in recent calibrations [61,70] or the use of too simple evolutionary models [71] may lead to altered results in dating analyses. Our approach included a calibration date in which coalescence should be negligible for usual population sizes in mammals [61] and we used a codon-partitioned model, which should avoid these problems. Nonetheless, an increase in the rate that we estimated for Galemys would only reduce, in the equivalent proportion, the split time of the mitochondrial lineages. Since our main purpose in this part of the work was to test if the separation of the mitochondrial lineages occurred in the Pleistocene, any increase in this rate would still support the Pleistocene split of the G. pyrenaicus mitochondrial lineages. Although introns were very useful for obtaining the Galemys-Desmana split time in the first step of our dating analysis, the low variability of these sequences (Table 2) did not allow us to use them in a multilocus dating analysis for the second step, which would have permitted a direct estimation of the population splits in a species tree framework. In fact, the lack of accumulated differences at the nuclear level between the four desman populations may indicate more recent separations than the ones indicated by the mitochondrial genes. Further studies with additional nuclear data will help to resolve these issues.
Pleistocene evolution of the Pyrenean desman populations
From the four inferred glacial refugia, other areas of the current distribution range would have been subsequently colonized during the Holocene, as depicted in a schematic scenario of the evolution of the desman populations (Figure 9). Given the strong geographic pattern of the four mitochondrial lineages of G. pyrenaicus, it is possible to speculate about the relative locations of the refugia where they evolved. Different independent pieces of data support that the NW of the Iberian Peninsula served as the major glacial refugium for the desman populations. In particular, we found the greatest genetic diversity of the species in the northwestern part of the Iberian Peninsula and much more homogeneous populations towards the eastern parts of the desman distribution. In addition, distribution models of the Pyrenean desman projected to the conditions of the LGM predicted the highest probabilities of potential presence in the same northwestern area. The coincidence between the highest genetic diversity and the predictions of potential presence in the LGM in the same area is very remarkable, but it is the expected result from a classical phylogeographic scenario in which only part of the genetic pool from the glacial refugia colonized new areas. Furthermore, a signal of population expansion was detected in the northwestern mitochondrial lineage. This area was a likely glacial refugium for other species that also depend on aquatic habitats such as the golden-striped salamander [72], so this region probably preserved optimal temperature and pluviometric conditions for species with these particular requirements during the successive glacial cycles.
An additional refugium situated in the Iberian Range or the Central System could have given rise to the populations of lineage A2, but the location of this refugium remains very speculative due to the scarcity of data for this lineage. Given the genetic proximity of this lineage to the northwestern A1 lineage, the refugium could have
been situated somewhere in the Central System rather than in the more distant Iberian Range. If this were the case, part of the Iberian Range (Cameros, Urbión and Cebollera Mountains) would have been recently colonized. However, the dispersal, at least of females, would not have progressed towards the northwestern parts of the Iberian Range (Demanda Mountains).
Lineage B1 could have evolved in a refugium in the Cantabrian Mountains, where the maximum genetic diversity for this lineage was found. From this refugium, the species would have colonized the northwestern part of the Iberian Range (Demanda Mountains). However, at least the females would not have continued the dispersal towards more southeastern parts of the Iberian Range (Cameros, Urbión and Cebollera Mountains). Dispersal of lineage B1 from its putative refugium towards the western parts of the Cantabrian Mountains would have also been limited, and, at least the females, would not have crossed to the areas occupied by lineage A1.
Despite potentially suitable refugia within the Pyrenees [73], the Pyrenean populations of desmans, which belong to lineage B2 and are highly homogeneous genetically, must have originated from a distant refugium after a severe bottleneck. This refugium could have been placed
towards the middle of the distribution this lineage, such as in the Basque Mountains, as previously suggested according to the current distribution of the species [74]. The colonization of the Pyrenees must have been quite recent and likely occurred very quickly and through an important bottleneck, as deduced from the very low nuclear and mitochondrial diversity observed in the desmans of these mountains. From this putative refugium, the populations of this lineage also dispersed in other directions, towards the areas currently occupied by lineage B1, so that there is currently mixing of both B lineages in some rivers. The areas proposed here as likely glacial refugia for lineages B1 and B2, in the Cantabrian Mountains and the Basque Mountains, respectively, have been previously postulated to maintain potential refugia for many other species associated to humid climates, including mammals [75,76] and plants [77].
The distribution of mitochondrial lineages and their variability allowed us to infer a clear postglacial expansion of the desman populations from the refugial areas. This range expansion should be in principle accompanied by an increase in population size, but the population expansion parameters of the mitochondrial sequences did not show statistically significant support in all lineages (Table 1). This is particularly noteworthy for the B2 lineage, in which the low genetic variability across the Pyrenees point to a strong bottleneck before the colonization of these mountains. However, the extremely low variability of this lineage (only four variable sites; Table 1) surely diminishes the statistical power of the expansion statistics. Additional mitochondrial data or more variable markers should be used to formally test the existence of these demographic expansions.
It is also worth noting that the range expansions hypothesized here could have taken place during particular periods of very favorable conditions, as shown for other species [78]. In the case of the Pyrenean desman, the abundance of humid habitats during the deglaciation periods could have helped to quickly colonize new areas and extend its distribution range from the glacial refugia.
Thus, the evolutionary history of the Pyrenean desman supports the “refugia within refugia” hypothesis [6], which highlights that the Iberian Peninsula and likely other South European peninsulas cannot be regarded as homogeneous refugia but rather as centers of development of multiple refugia that gave rise to distinct evolutionary lineages within many species. Our results further extend this hypothesis by showing that peninsulas would have helped to develop, not only complex isolation mechanisms, but also the whole glacial processes of contraction and dispersal, leaving strong footprints on the genetic structure of endemic species such as the Pyrenean desman. Although these clear genetic traces had been mostly identified in species of continental distribution [7], some of which left distinctive lineages [79] or even species [80,81] in the southern peninsulas, a growing number of endemic or semi-endemic species shows similarly complex population history patterns within the Iberian Peninsula [72,82-85]. We show here that the genetic structure of the Pyrenean desman, a highly specialized mammal, was also affected by the whole glacial processes at a peninsular scale. Small influence of the river network on the genetic structure of the Pyrenean desman Contrary to the expectation that the genetic structure of a species with a semi-aquatic lifestyle and a strong dependence for clean waters, such as the Pyrenean desman, would be highly related to rivers and drainage basins, we found that only a small proportion of its genetic variation can be attributed to the grouping of populations by major river systems. In fact, identical mitochondrial haplotypes can be found at both sides of different mountain ranges, explaining the lack of strong differentiation among basins. Thus, these data allow us to infer that gene flow between basins exists or existed in a not so distant past. In conclusion, the genetic structure of the Pyrenean desman has been more influenced by the history of the Pleistocene glaciations than by its current aquatic habitat distribution, in spite of the strong fragmentation of such specialized habitat. This situation is intermediate between strictly aquatic organisms, whose genetic diversity has been more conditioned by river basins [1], and highly mobile semi-aquatic mammals, such as the Eurasian otter, whose genetic diversity is totally unrelated to river basins [86].
Strong signatures of isolation in the contact zones
The most unexpected finding in the genetic structure of the Pyrenean desman was the existence of narrow contact zones between the mitochondrial lineages that came into secondary contact after the post-glacial recolonization, with no apparent mixing among them. Actually, dispersal of the four lineages in different directions from the peripheral glacial refugia and the lack of suitable areas in the central parts (Meseta Central) have created an interesting circular distribution of the Pyrenean desman (Figure 9). In addition, the interrupted dispersal of lineage B1 in both clockwise and anticlockwise directions, and of lineages A1 and A2 towards the areas of lineage B1, have created two prominent genetic gaps, that is, there are two replicate contact zones of the major genetic groups, A and B (Figure 9).
The strongest genetic gap was found in the middle of the Iberian Mountain Range, one of the places where the A and B groups meet (Figure 1A). The 23 samples collected in six rivers of the Iberian Range revealed that individuals belonging to both major lineages were present in this area. However, with the samples available so far, the lineages are segregated and have not been found together in any river stretch. In fact, we can trace a separation line (basically along the valley of the Najerilla river) that seems to restrict the dispersal of female desmans. The second genetic gap was found in the middle of the Cantabrian Mountains and it also affects the same major lineages, A and B. Despite conducting several surveys in this area of the Cantabrian Mountains, we could not get more samples to narrow the closest distance between both lineages. Therefore, we cannot determine at present whether or not some mixing of lineages occurs in this contact zone. However, the lack of penetration of females of one lineage into the distribution area of the other lineage is a remarkable circumstance in both contact zones, where no apparent barriers to dispersal of desmans exist. Although similar situations have been observed in other species [60,66,87-89], including some of the Iberian Peninsula [72,82-84,90,91], certain degree of permeability through the contact zones is normally observed in these species, in contrast with the more strict situation seen in the Pyrenean desman.
This phenomenon of competitive exclusion within species could have adaptive or neutral (demographic) causes [92]. Although adaptive processes cannot be excluded, it has been suggested that saturation of the habitat in the contact zones would inhibit female migration in some species (density blocking) [91,93]. This would explain why some of these species have dispersed hundreds of kilometers through empty spaces from glacial refugia but now seem unable to cross a stretch of a few kilometers [92].
The analysis of contact zones discussed so far has been based on mitochondrial data and therefore only refers to the dispersal pattern of females. Although obviously a crucial aspect of the species biology, it may not tell the whole story. In fact, in many species in which nuclear data was obtained, it has been observed that these barriers were not so strong or that they were absent for these markers, indicating male-biased dispersal [83,89,94]. Our intron sequences did not show enough variability within G. pyrenaicus to analyze these aspects in depth. However, three variants of the obtained SNPs (Figure 6) exhibited enough geographic extension to be useful in the analysis of dispersal [95]. The three derived mutations showed a contiguous distribution within the sampling localities, suggesting that they arose in place and are of recent origin. In fact, one of the three mutations (in intron DHRS3-3; Figure 6) crosses the Cantabrian Mountains contact zone, suggesting that some male-driven dispersal has occurred through it, giving rise to certain degree of introgression. However, additional nuclear genetic data will be necessary to study these aspects in a more quantitative manner. So far, radio-tracking and recapture data of desmans have not revealed sex-biased dispersal [25] but data are still very scarce. Therefore further studies, both genetic and behavioral, should be carried out to better understand the mobility patterns and barriers to dispersal of the Pyrenean desman.
Subspecies
The existence of two main mitochondrial groups in the Pyrenean desman could in principle correspond to the two described subspecies, G. p. pyrenaicus and G. p. rufulus, but the distribution of the mitochondrial groups does not perfectly fit with any of the proposed distribution areas for the two subspecies, which have been very unstable in previous works [19-21]. However, none of the previous studies trying to delimit morphologically the subspecies took into account the boundaries between the populations revealed in this work. They rather mixed specimens belonging to different mitochondrial lineages in the analyses. For example, all specimens of the Iberian Range were pooled into a single population when, in fact, there are two distinct lineages in this region. This could have hindered the detection of significant morphological differences between subspecies [19,21]. Future studies aimed at assessing the validity of these subspecies should analyze phenotypic differences between these groups and possible morphological gradients in the contact zones detected in this work. For the moment, according to the genetic results and the corresponding type localities of the subspecies [20], the populations of mitochondrial group A would correspond to subspecies G. p. rufulus, and those of group B to subspecies G. p. pyrenaicus.
Implications for conservation of the Pyrenean desman
The Pyrenean desman is legally protected in the four countries where it is present and it was classified as “Vulnerable” in the IUCN Red List [12]. In addition, the populations of the Central Mountain System, in the southern part of the distribution, were recently catalogued by the Spanish Government as “In danger of extinction”,
which is the highest protection category. The desman is therefore one of the most threatened mammals of the Iberian Peninsula and, by extension, of the European continent. Indeed, many data seem to indicate a substantial decline of the Pyrenean desman in the Central System in recent times [11]. Actually, our own surveys did not yield any desman excrement in several localities of the Central System where the species had been captured in the last few decades, which forced us to rely on museum samples for our DNA work. More targeted surveys in the most southern parts of the historical range will be of utmost importance in future demographic and genetic studies of the species.
The genetic diversity of the Pyrenean desman was very small in its whole range, as confirmed with both mitochondrial and nuclear markers. Regarding mitochondrial data, for which there are more data for comparison, the nucleotide diversity of the Pyrenean desman is around four times smaller than the mean for mammals [96], and it is particularly low in some areas such as the Pyrenees. Interestingly, however, the Pyrenean populations have been until recently in a relatively good state of conservation [18]. In fact, as we have shown, this low genetic variability was likely due to a recent colonization of the Pyrenees (and not necessarily to a decline of these populations). However, it is important to be aware of the populations with the lowest genetic diversity values in case of future unforeseen environmental changes, which might be more detrimental for them.
The conclusions about the lack of strong genetic differentiation among river basins of the Pyrenean desman may also have implications for conservation purposes. In particular, these results allow us to infer that desmans have not been confined to the river basins where they inhabit and that they can move, or have moved in the recent past, through at least some of the watersheds. Therefore, connectivity between some water basins should not be discarded, in certain cases, to prevent or to reverse an excessive fragmentation of the populations. However, future studies will be necessary to determine the amount of recent gene flow between specific basins in order to properly inform conservation actions in this regard.
A crucial aspect that should be certainly taken into account in conservation programs is the delimitation of G. pyrenaicus into the four mitochondrial lineages found in this work. These lineages started to diverge during the Pleistocene glaciations and, in consequence, their integrity should be preserved until further studies establish
the exact degree of genetic exchange between these populations [97]. Therefore, following a precautionary principle, these lineages should be considered as different evolutionary units for conservation purposes. In particular, great care should be exercised to avoid any translocation of individuals between these units and thus preserve both the integrity of the Pyrenean desman and its evolutionary history.
Conclusions
Mitochondrial and nuclear data in the Pyrenean desman (Galemys pyrenaicus) allowed us to study the phylogeography of this species and provided evidence for an evolutionary history deeply influenced by the Pleistocene glaciations. One of the most striking findings of this work was the existence of a strong phylogeographic structure in the Pyrenean desman, in which two large groups, A and B, were subdivided into two further groups to give a total of four mitochondrial lineages with parapatric distribution (A1, A2, B1 and B2). Two narrow contact zones between the major groups (A and B), one in the Iberian Range and the other in the Cantabrian Mountains, indicate incomplete mixing after the postglacial recolonization, at least for females. Nuclear data seem to indicate some degree of gene flow in these contact zones but more data will be necessary to further study the dispersal patterns of the desman. It is interesting to note that the presence of two major and parapatric mitochondrial groups parallels the existence of the two described subspecies, G. p. pyrenaicus and G. p. rufulus, whose distributions roughly correspond to groups B and A, respectively. A dating analysis of the desmanines allowed us to estimate that the separation of the major mitochondrial lineages likely occurred in the Middle Pleistocene. In addition, both the geographic variation of genetic diversity (with the populations of highest diversity in the NW part and those of lowest diversity in the Pyrenees) and a species distribution model projected to the LGM coincided in indicating that the most important glacial refugium was in the NW of the Iberian Peninsula. Other minor refugia can be postulated in other parts of the distribution areas of the present mitochondrial lineages. A Holocene expansion from these refugia, but interrupted at the contact zones, led to the current parapatric distribution of the mitochondrial lineages.
The Pyrenean desman is an endangered species and its situation has worsened during the last few years in part of its distribution range, particularly in the most southern populations. In order to undertake the most favorable actions for the long-term survival of this species, conservation programs should keep in mind the peculiar genetic patterns found in this work. Most importantly, artificial mixing of desmans and, particularly, of individuals belonging to different lineages should be avoided.
At the moment, almost no natural exchange between the lineages with different glacial origins has been observed and therefore no artificial translocations between them should be carried out until further studies establish the exact degree of genetic exchange between these populations. Although only following these criteria in management plans does not guarantee the conservation of the species, it would be essential to take this information into account in order to prevent an aggravation of the status of this singular species.
|
Como nenhum dos dois estudos inclui Galemys pyrenaicus da Serra da Estrela tem de ficar em aberto se a Serra da Estrela representa talvez um quinto centro de refúgio para esta espécie. No entanto, é mais provável que ramos de colonização dos centros refugiais, ou o da Galiza (A1) ou o do Sistema Central Ibérico (A2), chegaram à Serra da Estrela. Os Galemys pyrenaicus da Serra da Estrela pertencem assim provavelmente à subespécie rufulus o que indica também o seu tamanho (M. L. Madureira & M. G. Ramalhinho)[6].

Anotações e Bibliografia
[1] Council of Europe. SEMINAR ON THE BIOLOGY AND CONSERVATION OF EUROPEAN DESMANS AND WATER SHREWS (Galemys pyrenaicus, Desmana moschata, Neomys spp.) Ordesa, Spain, 7- 11 June 1995
[2] http://www.alchata.es/?cat=4&paged=8
[3] María José LÓPEZ-FUSTER, Rosa García-Perea, Rosa Fernández -Salvador, Julio Gisbert and Jacint Ventura: Craniometric variability of the Iberian desman, Galemys pyrenaicus (Mammalia: Erinaceomorpha: Talpidae). Folia Zool. – 55(1): 29–42 (2006)
[4] Javier Igea, Pere Aymerich, Angel Fernández-González, Jorge González-Esteban, Asunción Gómez, Rocío Alonso, Joaquim Gosálbez and Jose Castresana: Phylogeography and postglacial expansion of the endangered semi-aquatic mammal Galemys pyrenaicus. BMC Evolutionary Biology 13:115. 2013.
[6] M. L. Madureira & M. G. Ramalhinho: Notas sobre os Insectivora e Rodentia;a Portugueses 175
Sem comentários:
Enviar um comentário